
Mijn proefschrift uit 1987
voor Sophie
en Hieronymus
Men vraagt me waarom ik zoveel nummers wyd aan buitenissigheden. Eilieve, wanneer gy goederen te laden hebt in ’n vaartuig, en ge vindt dat vol, overvol … begint ge dan niet met lossen, reinigen, schoonvegen?
Is niet het wegruimen van ’t schadelyke, ’n even nuttige, ’n even noodzakelyke arbeid als ’t aanbrengen van het goede?
Multatuli, idee 178
ten geleide
Het boek dat nu voor u ligt is voor een proefschrift van een bioloog vrij ongebruikelijk van opzet. U zult er geen feitenkennis uit opdoen die niet ook al via eerdere publicaties van anderen beschikbaar was. Het gaat eerder over woorden dan over feiten. En wel over woorden die een belangrijke rol spelen in discussies over humane sociobiologie. Het is geschreven in de overtuiging dat een zinvolle gedachtenwisseling over de voors en tegens van een sociobiologische benadering van menselijk (en dierlijk!) gedrag alleen mogelijk is wanneer er voldoende helderheid bestaat over de betekenis van de beweringen van voor- en tegenstanders. En daarvoor is eerst en vooral begripsmatige helderheid nodig.
Dit boekje zou er niet gekomen zijn zonder hulp van anderen. Enkele van hen noem ik hier graag persoonlijk. De volgorde is vooral chronologisch.
Wim van der Steen heeft me van dag tot dag begeleid. En dat was geen makkelijke taak. Hij heeft me elk probleem waar ik overheen dreigde te stappen voorgehouden, en me steeds opnieuw geholpen oplossingen (als ’t moest: redactionele) te bedenken. Het duurde soms even voor ik daar blij mee was, en ik heb heel wat onvrede op hem afgereageerd, maar steeds als ik de moed kwijt was hielp hij me weer op weg.
In de eerste fase van mijn onderzoek had ik vaak geen idee waar het (en ik!) heen zou moeten. Avondenlange gesprekken met Jet Rootlieb verdreven de onrust daarover.
Professor Lever heeft mijn hele verhaal met zorg gelezen. Hij heeft me er niet van kunnen weerhouden in de eerste persoon enkelvoud te schrijven, maar zijn inhoudelijke commentaar heeft me steeds aan het denken gezet, en dat leidde vaak tot veranderingen.
Datzelfde geldt voor het commentaar van Peter Sloep. Ik waardeer het zeer dat hij daar, naast een nieuwe baby en een nieuwe baan, tijd voor heeft vrijgemaakt. Ik denk bovendien met plezier terug aan de gastvrijheid van Sioe Li en hem, zowel in Zuidbroek, als in Guelph en Schaesberg.
Nienke van der Baan was de enige niet-bioloog onder mijn eerste lezers. Zij bevestigde zowel mijn angst dat veel van wat ik zeg over vakken waar ik niet in thuis ben minder genuanceerd is als ik zou willen, als mijn hoop dat dit boek voor een ‘buitenstaander’ in principe te lezen is. Ik hoop dat haar kritiek me voor een onverantwoord gebrek aan nuance heeft kunnen behoeden.
Jan van Hooffs reactie stimuleerde me op de valreep nog tot tal van wijzigingen. Hij heeft mij daar twee zware maar leerzame middagen mee bereid.
Joop Vegter, ten slotte, nam de ondankbare taak op zich mijn hele boek na te vlooien op spel- en verwijsfouten.
Hen allen ben ik dankbaar voor steun en hulp. Maar vooral dank ik Sophie, die me er steeds weer aan herinnerde wat echt telt in het leven.
Amsterdam, 1987
Bart Voorzanger
stellingen
1. Onstelligheid is nozel.
2. Wie meent dat de beste beschikbare theorie beter is dan geen theorie gaat voorbij aan de heilzaamheid van stilte.
3. Hoewel beoefenaren van de natuurwetenschap zich graag beroepen op hun empirie en de verifieerbaarheid van hun uitspraken, hebben zij elk voor zich het overgrote deel van hun ‘kennis’ van horen zeggen. Enige bescheidenheid tegenover andere gelovigen zou hen dus niet misstaan.
4. Religiositeit begint bij het inzicht dat onbeantwoordbare vragen belangrijk zijn.
5. Wie uit het succes van de moderne techniek de juistheid van moderne natuurwetenschappelijke theorieën afleidt, veronachtzaamt de mogelijkheid dat beide slechts samengaan doordat ze voortkomen uit eenzelfde bron, en zet te snel de stap van correlatie naar oorzakelijk verband.
6. De redenering dat een wetenschapsbeoefenaar niet verantwoordelijk is voor misbruik van door hem ontwikkelde technieken omdat we immers ook een hamerfabrikant niet verantwoordelijk houden voor een moord begaan met een door hem gemaakte hamer, gaat in één opzicht mank: een moord begaan met een hamer wordt maar zelden uitgelokt door gestook van een hamerfabrikant in een van oorsprong harmonieuze relatie.
7. De veel gehoorde stelling dat wij ethisch (nog) met toe zijn aan de mogelijkheden van de moderne techniek suggereert ten onrechte dat wij ethisch wel zijn toegerust voor mogelijkheden die minder heet van de naald zijn, en verwart derhalve een kwestie van ethiek met een kwestie van wennen.
8. Er zal altijd een doodsoorzaak nummer één zijn.
9. Volgens Van Duivenbode snappen veel leken niet hoe natuurlijke selectie werkt doordat biologen bet begrip adaptatie verkeerd gebruiken. Met het woordgebruik in haar eerste stelling – ‘De voortplantingsstrategie van de hermafrodiete slak Lymnaea stagnalis is erop gericht dat zelfbevruchting zoveel mogelijk wordt beperkt.’ (cursivering, afgezien van de soortnaam, van mij) – werkt ook zij het onbegrip van leken in de hand.
(Y.A. van Duivenbode. 1984. Sexual behaviour of the hermaphrodite freshwater snail Lymnaea stagnalis. Proefschrift vu, stellingen 1 en IX.)
10. Kuipers’ explicatie van de functionele benadering in de biologie laat een aantal dingen zien:
(1) dat uitspraken die aan bepaalde kenmerken van bepaalde organismen een functie toeschrijven iets betekenen, en dat zulke uitspraken een rol spelen in functionele analyses,
(2) dat er in de biologie, náást wat biologen een functionele verklaring noemen, ruimte is voor argumenten die wel deductief geldig zijn,
(3) dat functionele uitspraken na logische reconstructie van onderzoekspraktijken evenmin een goede basis zijn voor het deductief afleiden van kenmerksbeschrijvingen als na logische reconstructie van verklaringen.
De auteurs die Kuypers kritiseert zullen met (1) en (2) geen moeite hebben, en (3) laat zien dat hij er niet in slaagt zijn hoofdbezwaar tegen de visie van Hempel te omzeilen. De meeste bezwaren die Kuypers inbrengt tegen eerdere reconstructies van de zogenaamde functionele verklaring zijn dan ook onverkort van toepassing op zijn eigen reconstructie.
(Th. A. F. Kuipers. 1986. The logic of functional explanation in biology. In: The task of contemporary philosophy (Proceedings of the 10th International Wittgenstein Symposium) blz. 110-114. Hölder. Pichler & Tempsky, Wenen.)
11. ‘Inclusive fitness’ is een overbodig begrip.
12. Inspanningen om tweede-geldstroomgeld te verwerven scherpen het vermogen succesvolle subsidieaanvragen te schrijven méér dan het vermogen zinvol wetenschappelijk onderzoek te doen.
13. Arbeidsmobiliteit en sociale desintegratie zijn onlosmakelijk met elkaar verbonden.
14. Genetici die speculeren over de mogelijkheid om eigenschappen van het ene organisme middels DNA-recombinanttechnieken over te planten in andere organismen zijn uitsluitend in overdrachtelijke zin bezig met het overplanten van eigenschappen: ze naaien hun publiek een oor aan.
15. Gezien de enorme verwachtingen die men van haar mogelijkheden heeft is het raar dat men de biotechnologie niet allereerst heeft ingezet voor de productie van Haarlemmerolie.
1. inleiding
In 1975 publiceerde Edward O. Wilson een boek dat de geboorte van een nieuwe biologische discipline inluidde en dat zeer veel stof zou doen opwaaien: Sociobiology, the New Synthesis.[1] Over dat boek zal ik het maar zeer terloops hebben, over wat er volgde des te meer. Dat namelijk vormt de aanleiding tot mijn onderzoek.
1.1. sociobiologie
Wat sociobiologie precies is is minder duidelijk dan men verwachten zou bij een vak dat zo duidelijk bij één boek begint. Dat ligt zowel aan Wilson als aan wat er verder allemaal gebeurde. Het was duidelijk Wilsons bedoeling een samenhangende theorie te ontwikkelen die een evolutionaire verklaring biedt voor alles wat zich aan sociale interacties tussen dieren voordoet. (Wat dat is, een evolutionaire verklaring, komt in hoofdstuk 5 aan de orde.) Nu is Wilson een omnivoor. Alles wat er vóór hem aan hypotheses over de evolutie van sociaal gedrag bijeengedacht was plus een flink deel van wat er aan praktisch onderzoek was verricht komt in zijn boek samen. Het is daardoor een weinig kritisch en slecht verteerd allegaartje geworden.[2] Bovendien wordt het boek in veler ogen ontsierd door twee onvoorzichtige hoofdstukken: het eerste, waarin de plaats van het vak temidden van andere vakken in wel erg zelfverzekerde termen wordt geschetst en het laatste, waarin de evolutie van de mens wordt behandeld.
Wilson heeft een vrij eenzijdig mensbeeld en hij toont zich wars van menswetenschappelijke inzichten. Naar Wilsons mening zullen we ’s mensen doen en laten vooral moeten verklaren vanuit de overlevings- en voortplantingskansen die dat doen en laten biedt. Elke andere interpretatie is onwetenschappelijk.
Veel mensen geloven in goden en in hoge waarden. Je zou dat kunnen verklaren door aan te nemen dat die goden en waarden bestaan, en dat de mens daar weet van heeft. Wilson wijst dat beslist af. Voor hem is al dat geloof niet meer dan functioneel zelfbedrog waaraan wij ons vermogen danken om te leven in succesvolle groepen die door een gedeelde overtuiging bijeen worden gehouden. Maar bedrog blijft het.[3] En op eenzelfde oneerbiedige manier schrijft hij over de verhouding tussen de seksen, en tussen rassen. Veel van wat welwillende westerlingen in die verhoudingen stoort wordt door hem bovendien verklaard op een manier die sommigen haast als een rechtvaardiging ervaren. Ongelijkheid en vreemdelingenhaat zijn een natuurnoodzakelijkheid waarzonder wij niet hadden bestaan en die zo diep in ons wezen verankerd ligt dat we er niet makkelijk van af zullen komen. Of Wilson dat als rechtvaardiging bedoelt blijft duister, maar het laat zich zeker zo lezen.
Wilsons verklaring voor het ontstaan van religie en ethiek heeft, zo op het eerste gezicht, haar concurrenten. Antropologen, sociologen en theologen, gelovige en ongelovige, hebben allerlei andere opvattingen over ontstaan en functie van godsdiensten en moraal. Nu hoeven al die verklaringen niet met die van Wilson te botsen,[4] maar Wilson was (in elk geval in 1975) van mening dat ze dat wel degelijk doen. De beoefenaren van de menswetenschappen die religies en ethische systemen vaak anders interpreteren dan Wilson zullen zijns inziens door de evolutiebiologie tot inzicht moeten worden gebracht.
Kortom, de sociobiologie legt in Wilsons optiek de basis voor een nieuw, en nu wel wetenschappelijk, inzicht in de menselijke natuur.[5] In een volgend boek, On Human Nature uit 1978, heeft hij zijn ideeën over de mens voor een breed publiek uitgewerkt.
Als ik het in het vervolg over sociobiologie heb bedoel ik daarmee in principe het deelgebied van de biologie waarin de evolutie van – met name sociaal – gedrag centraal staat. In die omschrijving zitten een paar elementen waar ik iets meer over zeggen wil. Sociobiologie is een vak, en wat mij betreft wordt een vak vooral gekenmerkt door het soort vragen dat zijn beoefenaren stellen, en niet bijvoorbeeld door één bepaalde theorie, één bepaald verklaringsschema. Je kunt dus sociobiologie bedrijven zonder de ideeën van een bepaalde sociobioloog te onderschrijven. Dat klinkt misschien wat triviaal, maar in dit geval is het dat toch niet helemaal. Ik vind sociobiologie een vak met bestaansrecht. Als ik dat zeg tegen een verklaarde tegenstand(st)er van Wilsons verhalen over mensengedrag leidt dat soms tot verontwaardiging: ‘Dus jij vindt dat Wilson gelijk heeft met zijn racistische en seksistische onzin!’ Welnu, dat vind ik niet, maar ik vind wel dat de vragen die sociobiologen stellen gesteld mogen worden. En ik denk dat er weloverwogener en minder bevooroordeelde antwoorden mogelijk zijn. Er zijn ook feministische sociobiologen, om maar iets te noemen. Die komen we in hoofdstuk 5 nog tegen. En er zijn soms belangrijke verschillen in opvatting tússen de sociobiologen die in veel kritieken wat erg snel op één hoop worden gegooid. Richard Dawkins, bijvoorbeeld, is regelmatig het slachtoffer van die onzorgvuldigheid.[6] Nu geeft hij daar wel aanleiding toe door soms wat controversiële voorbeelden te gebruiken. In een van zijn boeken illustreert hij de betekenis van het begrip ‘genetische bepaaldheid’ met een voorbeeld waarin hij aanneemt dat het verschil in breivaardigheid tussen jongetjes en meisjes genetisch bepaald is. En dat zal menig feminist de haren te berge doen rijzen. Maar het gaat in dit soort gevallen altijd om hypothetische voorbeelden en voor de goede verstaander is dat uit de context ook duidelijk. Dawkins’ theorieën gaan niet over menselijk gedrag, zoals die van Wilson, maar over het precieze mechanisme van natuurlijke selectie (zie hoofdstuk 3). En de rest is grappenmakerij.[7]
Sociobiologie gaat over evolutie. Maar daarmee is nog weinig gezegd. Dé evolutietheorie bestaat niet. Evolutiebiologie is een gebied vol boeiende problemen.[8] Sociobiologen zijn meestal sterk geneigd bij evolutie meteen aan natuurlijke selectie en dus aan genetische factoren te denken. We zullen in hoofdstuk 2 zien dat dat erg eenzijdig is. Over de mechanismen die verantwoordelijk zijn voor evolutionaire veranderingen is het laatste woord nog lang niet gezegd. Maar evolutiebiologie is meer dan een theorie over mechanismen. Voor een goed evolutionair begrip van de eigenschappen van een bepaald organisme is ook enig inzicht nodig in de historische ontwikkeling die tot die kenmerken leidde. De sociobioloog zal dus een beeld moeten krijgen van de historische[9] ontwikkelingen van sociale systemen en gedragingen. Ik zal later, aan de hand van voorbeelden, laten zien dat een voldoende betrouwbare geschiedenis van onze soort op dit moment nauwelijks geschreven kan worden[10] (zie hoofdstuk 5).
Hiermee is een belangrijke groep van de problemen die de aanleiding voor mijn onderzoek vormden ruw gekarakteriseerd. Het wordt nu tijd om iets meer te zeggen over de manier waarop ik te werk ben gegaan. Ik heb dit onderzoek niet gedaan omdat ik buitensporig geïnteresseerd ben in, al dan niet menselijk, sociaal gedrag. Mijn eerste belangstelling gaat uit naar de rol van taalproblemen bij het ontstaan van misverstanden en meningsverschillen over wetenschap. En ik denk dat het zichtbaar maken van die taalproblemen kan bijdragen tot het oplossen van onenigheid.
1.2. meningsverschillen en begripsverwarring
Meningsverschillen over wetenschap hebben maar zelden één simpele oorzaak. Aan de discussie over sociobiologie is dat duidelijk te zien. De deelnemers aan discussies over sociobiologie verschillen al van mening over feitelijke zaken. Over de vraag bijvoorbeeld in hoeverre gedragsverschillen tussen mannen en vrouwen genetisch bepaald zijn. Sommige sociobiologen menen dat het belang van genen hier vrij groot is, sommigen van hun tegenstanders achten het heel klein. Wie er gelijk heeft weten we niet. Voor we dat weten zal er nog heel wat onderzoek gedaan moeten worden.
Voor- en tegenstanders zijn het ook oneens over het beláng van de biologische kennis die sociobiologen aandragen. Stel dat het gedragsverschil tussen vrouwen en mannen in belangrijke mate genetisch bepaald is, moeten we dan daarom vrede hebben met dat verschil, of moeten we juist extra hard vechten voor een wereld waarin die verschillen zo klein mogelijk zullen zijn? Er zijn sociobiologen[11] die neigen tot het eerste standpunt, ze vinden dat realistisch. En hun tegenstanders zullen zeker niet ontkennen dat het goed is de wereld te kennen, maar ze vinden niet dat je je door die kennis moet laten voorschrijven welke idealen je nastreeft. Bovendien hebben ze soms hun twijfels waar het de status van wetenschappelijke kennis betreft.
Achter elke wetenschappelijke uitspraak schuilen enorm veel onbewezen veronderstellingen. Je kunt geen natuurwetenschap bedrijven als je niet beschikt over een flink vertrouwen in eigen waarnemingen, als je niet gelooft dat de dingen die je bestudeert kunnen worden beschreven in termen van materiële eenheden en ketens van oorzaken en gevolgen. Sociobiologen vertrouwen hun waarnemingsvermogen, en geloven in een door oorzaken geregeerde materiële wereld, ook waar het het gedrag van mensen betreft. Hun tegenstanders vaak niet.
Verschillen van mening over wetenschapsfilosofische, ideologische en empirische kwesties spelen dus een grote rol in discussies over sociobiologie. Over empirische kwesties zal ik het hier niet hebben. Dit is een theoretische verhandeling. Maar ook de ideologische en wetenschapsfilosofische vragen komen nauwelijks aan bod. Ik vind ze heel interessant, maar ik zal er weinig over zeggen. Ik zal alleen kort maar met veel plezier mijn eigen wetenschapsbeeld onder woorden brengen. Ik doe dat vooral omdat ik hoop dat dat mijn benadering begrijpelijker maakt. Daarna zal ik schetsen wat ik verder wél zal doen, en waarom dat nuttig zou kunnen zijn.
1.2.1. kennis – een intermezzo
De mens leeft in een baaierd van indrukken en het lukt hem slecht die over zich heen te laten gaan. Daarom vertelt hij. Hij vertelt verhalen om indrukken te ordenen en de chaos te bezweren. Zou hij dat niet doen, zou hij zwijgen en al die indrukken alleen maar ondergaan, dan zou hij volgens sommigen de stem van de Schepper horen. Maar liever hoort de mens zichzelf.
Ik hecht eraan alle verhalen die worden verteld – van de Tau Te Tsjing en de Bijbel, via de Sprookjes van Moeder de Gans, tot Darwins Origin of Species – kennis te noemen. Kennis is geordende ervaring. Dat zal niet iedereen met mij eens zijn. Veel mensen noemen alleen hun eigen favoriete verhaal ‘kennis’ en geven daarmee een beperkte verzameling ervaringen en een bepaalde manier van ordenen een bevoorrechte positie.
De mens ordent ervaringen door ze in één groot verband te zetten, en een deel van de verhalendiversiteit komt voort uit de grote verscheidenheid aan verbanden die mensen hebben bedacht. Voor sommigen is de wereld een samenspel van oorzaken en gevolgen, voor anderen een speelveld van goden, en er zijn er die de wereld zien als een geheel van betekenisvolle, naar elkaar verwijzende, gebeurtenissen. Voor de eersten wast het water omdat de sneeuw smelt, voor anderen stroomt de rivier over vanwege een Gode onwelgevallige manier van leven, en voor de laatsten (in mijn onvolledige en vertekende opsomming) verwijst al dat water naar de grote cirkelgang der natuur (ik doe maar een gooi). De mens kiest niet alleen een bepaald soort verband als meest gezaghebbend, hij kiest ook voor bepaalde ervaringen – die hij dan bijvoorbeeld ‘feiten’ noemt – en negeert omzichtig heel wat andere. Dat wat ik kennis noem zegt net zo goed iets over de kenner, en zijn angst voor chaos, als over het gekende.
Vaak doen mensen een poging om hun eigen keus voor een bepaald verband en voor bepaalde ‘feiten’ te rechtvaardigen. Ze ontkennen zin en voordeel van andermans keuze, en benadrukken de zegeningen van hun eigen opvatting. Maar een rechtvaardiging die me overtuigde kwam ik nog niet tegen. Het ligt, geloof ik, niet zo in mijn aard om snel overtuigd te raken van de juistheid van een bepaalde opvatting en me te scharen in de rangen van haar aanhangers. Dat betekent niet dat ik een bepaalde visie niet serieus neem, maar op de een of andere manier struikel ik makkelijker over de beperkingen dan dat ik me optrek aan de geboden inzichten.
Natuurwetenschap[12] is in onze wereld een zeer prominente kennisbron. Net als alle andere heeft ook zij haar eigen wijze om eigen onovertroffenheid te beargumenteren. Allereerst laat zij zich voorstaan op objectiviteit en een zorgvuldige empirie.[13] Wetenschap gaat volgens haar bevlogen beoefenaars niet zo maar over indrukken en ervaringen in het wilde weg, maar over zorgvuldig waargenomen feiten. In de praktijk betekent dat dat de aspirant-onderzoeker eerst moet leren wat hij geacht wordt waar te nemen. Op basis van die selectief waargenomen feiten stellen onderzoekers theorieën op. Die worden getoetst aan nieuwe feiten. Net zo goed als nieuwe feiten worden getoetst aan de theorie. Het loopt dus al met al nog wel los met die harde empirische basis.
Wetenschapsbeoefenaars laten zich niet zelden voorstaan op de helderheid, de logica, de rationaliteit, en de verklarende kracht van hun bezigheden en producten.[14] En dat is niet helemaal terecht. Zij zouden vrij zwijgzaam zijn als ze louter logisch te werk gingen. Bovendien zijn centrale begrippen in de wetenschapstaal, ‘oorzaak’, ‘gevolg’, ‘gelijktijdigheid’, bijvoorbeeld, net zo vaag, diepzinnig en ondefinieerbaar als centrale begrippen in andere verhalenstelsels. En wie heeft ooit atomen gezien? Begrijp me goed, dit zijn geen verwijten, maar wel redenen voor bescheidenheid.
Kortom, zowel de manier waarop het uitgangsmateriaal (de ‘feiten’) wordt verzameld, als de ordeningsprincipes, ‘oorzakelijkheid’ en ‘logische samenhang’, rechtvaardigen een gezonde scepsis tegenover de (natuur)wetenschap. Wie zijn doelstellingen en praktische beslissingen uitsluitend probeert te baseren op wetenschappelijke overwegingen gaat niet zo vreselijk veel verstandiger te werk als iemand die zich laat leiden door het patroon van gevallen lotusstengels, of door de gedachten die na stil gebed bij hem bovenkomen.
Het zal u na deze ontboezemingen over mijn wetenschapsbeeld niet verbazen dat ik me niet geroepen voel partij te kiezen in discussies over sociobiologie. Ik voel me vaak vooral een verbaasde buitenstaander. Maar soms zie je vanuit zo’n positie dingen die de discussianten ontgaan. En wat me, naast de schrille klanken van gekwetste stemmen, vooral opvalt is de gigantische begripsverwarring. Daar zal het hier dan ook allereerst om gaan.
Ik zei al dat de meningsverschillen rond de sociobiologie veel verschillende oorzaken hebben. Misschien is begripsverwarring daarvan bij lange na niet de belangrijkste. Maar om te kunnen overzien welke reële problemen er nu precies spelen komt enige helderheid wel van pas.
1.2.2. methodologie
Ik zal in de nu volgende paragrafen iets meer zeggen over de rol en de status van de methodologie. Een waarschuwing vooraf. Wat ik hier over methodologie zeg heeft vooral betrekking op de situatie in de biologie, en niet op die in andere vakken. In veel sociale wetenschappen is expliciete aandacht voor methodologie min of meer vanzelfsprekend. In de biologie ligt dat vaak beduidend anders.
Het nuttig product van goed wetenschappelijk werk is een helder verhaal dat een nieuw licht werpt op de zaken waarover het gaat. Zo’n verhaal bestaat uit uitspraken die elk op zich ook helder moeten zijn. En, voor alle duidelijkheid, hier zal elke onderzoeker het mee eens zijn. Alleen, wetenschapsbeoefenaars beschikken niet altijd over alle technische vaardigheden die nodig zijn om onhelderheid te herkennen en op te lossen. Wetenschapsfilosofen, en meer in het bijzonder methodologen, zijn daar soms beter in.
De methodoloog werkt met een geordend stelsel van normen,[15] methodologische criteria, waaraan (stelsels van) wetenschappelijke uitspraken moeten voldoen.[16] Helderheid is een van die criteria. Daarover nu iets meer. Uitspraken kunnen alleen helder zijn als ze vervat zijn in woorden met een duidelijke betekenis. Die woorden mogen niet te vaag zijn. En ze moeten één betekenis hebben, ze mogen niet meerzinnig[17] zijn.
Een woord is vaag wanneer van allerlei dingen niet duidelijk is of die wel of niet met dat woord worden aangeduid. ‘Werk’ is een schoolvoorbeeld van een vage term. Sommige mensen maken onderscheid tussen werk en de dingen die ze thuis doen. Huishoudelijk werk is volgens hen dus geen werk. Gevraagd naar een preciezere omschrijving van wat werk is komen ze bijvoorbeeld met de volgende omschrijving: werk is iets waar je moe van wordt en dat je een inkomen oplevert. Het vermoeiende rondhangen in wachtkamers van uitkerende instanties afgewisseld met vruchteloos gesolliciteer zou dan werk zijn, en dat zal niet iedereen zo zien. Misschien moet de omschrijving worden uitgebreid: werk is iets waar je moe van wordt en dat een zinvol product plus een inkomen oplevert. Maar dat zou dan weer betekenen dat de producenten van elektrische tandenborstels en andere hebbedingen geen werk hebben, en dat zal niemand echt vinden. Dus rondom een vaste kern van wat iedereen werk noemt – dat wat de vuilnisman doet als hij vuil ophaalt en wat de tandarts doet als hij boort – ligt een ruim gebied van activiteiten die we soms wel en soms niet ‘werk’ zullen noemen. Die vaagheid maakt discussies over werk soms nogal moeizaam.
Een topsporter die een meester op het paard wordt genoemd kan zowel een begenadigd ruiter als een voortreffelijk turner zijn. ‘Paard’ is een meerzinnig begrip, het heeft een aantal totaal verschillende betekenissen. Dat zal niet vaak tot verwarring leiden, meestal blijkt uit de context wel om wat voor paard het gaat. Wat moeilijker ligt dat bij een term die hier later uitgebreid aan de orde komt. Een ‘altruïst’ wordt door sommigen omschreven als iemand die iets voor anderen over heeft, maar soms ook als iemand wiens gedrag hemzelf benadeelt terwijl het voor een ander voordelig is. Maar iets voor anderen over hebben en ze feitelijk bevoordelen hoeven niet samen te gaan. Wie zijn broekzakken niet onderhoudt en daardoor regelmatig geld verliest is wel een altruïst in de tweede maar niet noodzakelijk ook in de eerste zin van het woord.[18] Deze meerzinnigheid, zo zullen wij later zien, leidt ertoe dat sommige sociobiologen hun vak relevanter achten dan het zijn kan.
Wetenschappelijke begrippen zijn soms behoorlijk ingewikkeld. Dat hoeft op zich niet tot onhelderheid te leiden, maar doet dat in praktijk wel wanneer de ingewikkeldheid niet wordt gezien. Dat risico is vooral groot bij een bepaald soort ingewikkeldheid, die ik meerplaatsigheid zal noemen. Een begrip is meerplaatsig[19] wanneer het een verband legt tussen twee of meer dingen. ‘Groter zijn dan’ is een voorbeeld van een tweeplaatsig begrip. Het is een begrip met twee open plekken die moeten worden gevuld voor er iets betekenisvols ontstaat. ‘Groter zijn dan’ legt een verband tussen twee dingen: ding 1 is groter dan ding 2. En ‘staan tussen’ is drieplaatsig: ding 1 staat tussen ding 2 en ding 3. Zinnen als ‘A is groter dan’ en ‘A staat tussen B’ herkennen we snel als onvolledig. We zien kennelijk meteen hoeveelplaatsig de betrokken begrippen zijn. Maar soms gaat het mis. Mensen kunnen soms lang kibbelen over de vraag of Amsterdam nu groot is of niet. Meestal worden ze het er na een poosje over eens dat Amsterdam naar Nederlandse maatstaven een grote stad is, maar naar bijvoorbeeld Amerikaanse maatstaven niet. Indirect onderkennen ze daarmee dat ‘groot zijn’ een tweeplaatsig begrip is dat ze als eenplaatsig begrip gebruikten. En dat soort gebruik kan tot flinke verwarring leiden. In discussies over sociobiologie wordt ‘altruïstisch’ vaak als een- of tweeplaatsig begrip gebruikt, terwijl het, bij een bepaalde uitwerking, vier- of zelfs vijfplaatsig is (zie 3.2.2.2).
Het is ongetwijfeld overdreven, hoewel voor mij soms wel verleidelijk, om te stellen dat de meeste problemen in de wetenschap (en daarbuiten?) voortkomen uit onvoldoende aandacht voor begripsvorming. Maar ik ben ervan overtuigd dat begripsverwarring een belangrijke bron van wetenschappelijke onenigheid is.
1.2.2.1. methodologie: wetenschappelijk belang
Het soort onderzoek waar dit boek over gaat wordt niet altijd met evenveel enthousiasme begroet. Methodologen hebben duidelijk belangstelling voor stukken wetenschap die niet helemaal pluis zijn[20] en het is verleidelijk om die belangstelling ziekelijk te noemen. Aangemeten is dat oordeel niet. Je zou met hetzelfde recht de klinisch psycholoog kunnen verwijten dat hij een morbide belangstelling voor ongelukkige mensen heeft, of de bedrijfskundig adviseur dat hij altijd weer daar te vinden is waar zaken in het slop raakten.
Methodologen leveren vooral kritiek. Aan opbouwende adviezen en voorstellen voor een helderder theorie komen ze vaak niet toe, laat staan aan praktisch onderzoek dat wel door hun eigen beugel kan. Zou je als methodoloog niet beter zelf empirisch onderzoek kunnen doen? Ik denk het niet. Een goede methodoloog is niet ook vanzelfsprekend een goede praktisch onderzoeker, net zo min als een capabel ethicus vanzelfsprekend een goed mens, of een ervaren psycholoog vanzelfsprekend een toonbeeld van evenwichtigheid is. Bij empirisch wetenschappelijk onderzoek komt meer kijken dan helderheid en logische samenhang. Inventiviteit, handvaardigheid, en improvisatietalent zijn onontbeerlijk bij praktisch wetenschappelijk werk, en dat zijn geen eigenschappen waar een methodoloog over hoeft te beschikken.[21] We hebben nu eenmaal niet allemaal dezelfde talenten.
Natuurlijk mag je van een methodoloog verwachten dat hij bij al zijn kritiek ook vertelt hoe men het beter zou kunnen doen. En dat kan ook.[22] Maar het heeft weinig zin om adviezen te geven als daar niet om gevraagd is, en wat dat betreft is de relatie tussen biologen en methodologen vaak minder goed dan hij zou kunnen zijn. Hoe dat komt weet ik niet. Misschien stelt de methodoloog zich wel te veel op als schoolmeester, in plaats van als collega die nog heel veel leren moet. En misschien hebben biologen die praktisch onderzoek doen net iets te weinig oog voor methodologische valkuilen. Hoe dan ook, tot vruchtbare uitwisselingen komt het nog niet vaak. En dat is jammer.
Wetenschap bedrijven is meer dan om je heen kijken en opschrijven wat je ervaart. Dat zagen we al. Wetenschappelijke kennis wordt geacht op een specifieke manier bijzonder te zijn. Ze zou afstandelijk en algemeen zijn. Ze zou ons inzicht geven in de oorzakelijke samenhang der dingen. Ze zou ons in staat stellen om in allerlei processen in te grijpen en te voorzien wat de gevolgen van dat ingrijpen zullen zijn. En ze heet helder, ondubbelzinnig en voor iedereen toegankelijk en controleerbaar. Allemaal kenmerken die worden gebruikt om te rechtvaardigen dat wetenschappelijke kennis in onze samenleving een bijzondere plaats inneemt. Zo bijzonder dat zowel heel praktische beleidsproblemen als, in toenemende mate, ook meer fundamentele levensvragen aan de wetenschapsbeoefenaar worden voorgelegd, en niet aan de ziener of de dichter.
Ik zei al dat ik die bevoorrechte positie onwenselijk vind. Maar dat neemt niet weg dat wetenschappelijke verhalen naast allerlei andere hun bestaansrecht hebben. En ik denk dat wie op een gegeven moment benieuwd is naar wat de wetenschap ergens over te melden heeft erbij gebaat is dat het verhaal dat hij voorgeschoteld krijgt ook op de voor wetenschap kenmerkende manier tot stand kwam. Als het goed is, zijn methodologische criteria niet meer dan een uitwerking van die manier. Dat betekent dat een wetenschapper die wordt geconfronteerd met methodologische kritiek vier mogelijkheden heeft:
• hij kan laten zien dat de critici zijn werk niet goed hebben bestudeerd
• hij kan laten zien dat er iets mis is met de criteria waaraan zijn werk wordt getoetst
• hij kan toegeven dat de kritiek terecht zou zijn als zijn werk wetenschap was, maar die pretentie bij nader inzien laten vallen en een ander etiket op zijn verhalen plakken, of
• hij kan de kritiek accepteren en zich bereid tonen zijn werk te herzien.
Alleen in het laatste geval heeft het zin om te gaan praten over een andere opzet van het onderzoek.
In zekere zin is methodologie niet meer dan een spiegel die de onderzoeker zichzelf bij tijd en wijle kan voorhouden. Als hij daarbij schrikt van wat hij ziet, kan hij met behulp van de spiegel proberen het geziene te fatsoeneren. Samenwerking is dan het parool. Maar het heeft weinig zin de spiegel te verwijten dat die niet kant en klaar een verbeterd voorkomen levert.
Nog één opmerking tot slot van deze paragraaf. Als methodologische criteria een uitwerking vormen van wat voor wetenschap kenmerkend is, en als het zo moeilijk is om te voldoen aan die criteria, dan ligt het voor de hand dat een deel van wat wetenschap genoemd wordt dat in werkelijkheid niet is. Er zijn allerlei zaken waarover we (nog?) geen wetenschappelijke kennis bezitten. Maar dat betekent niet dat onderzoekers daarover zwijgen. Ze zullen er soms, eventueel onder druk, en als dat in hun aard ligt ook zeer voorzichtig, voorlopige uitspraken over doen. Het gaat dan niet langer om de resultaten van onderzoek, maar om de mening van onderzoekers, van ‘deskundigen’. Dus als ik van een verhaal weet dat het uit de mond van een wetenschapsbeoefenaar komt, weet ik niet genoeg. Het kan tot stand zijn gekomen na zeer zorgvuldig onderzoek dat zo veel mogelijk aan methodologische criteria voldoet, maar het kan ook niet meer zijn dan de mening van een deskundige. Dat wil ik dan wel graag weten. In het ene geval betekent het iets anders voor me dan in het andere geval. Daarom zou ik het woord ‘wetenschap’ liefst reserveren voor een relatief[23] beperkt geheel van activiteiten.
1.2.2.2. methodologie: maatschappelijk belang
Methodologie is niet alleen van belang voor onderzoekers. Er zijn meer betrokkenen. De lotgevallen van de sociobiologie illustreren dat. Sociobiologie is niet zo maar een nieuwe biologische discipline. Het is een vak waar zeer veel over geschreven wordt in de publiekspers: kranten, week- en maandbladen. Die publieke belangstelling heeft veel te maken met de eerder gesignaleerde pretenties van sommige sociobiologen dat ze iets wezenlijks over ’s mensen aard te melden hebben. Het mensbeeld van de sociobiologie was dan ook het centrale thema van veel boze reacties. De discussies die daaruit voortkwamen namen al snel een stereotype wending. Het veld raakte verdeeld in voor- en tegenstanders. De tegenstanders betoogden dat sociobiologie een verkapte poging was om een zeer bepaalde ideologie als wetenschap te presenteren en zo respectabel te maken. De ideologie in kwestie is dan de visie dat mensen met name naar sekse en ras ongelijk zijn en op grond van hun ongelijkheid ook verschillend behandeld moeten worden. De voorstanders stelden daar tegenover dat zij in het geheel niet van die ongelijkheid uitgaan, maar dat zij op grond van zorgvuldig wetenschappelijk onderzoek vermoeden dat met name het verschil tussen de seksen (over rassen spreken zij in feite niet of nauwelijks) biologisch verankerd is, en soms groter is dan men mocht hopen. Ik zal hier geen poging doen ‘het’ sociobiologiedebat uitgebreid weer te geven.[24] Het was en is verward en geëmotioneerd. Die verwarring wordt door niets gerechtvaardigd, de emoties eigenlijk alleen door het besef dat er belangrijke zaken op het spel staan.
Een centraal verwijt aan het adres van de sociobiologen is dat zij allerlei fascistoïde bewegingen in de kaart spelen door deze aan schijnbaar wetenschappelijke argumenten voor hun racistische en seksistische ideeën te helpen. Dit verwijt wordt gestaafd met citaten uit de partijbladen van deze bewegingen, waarin naar sociobiologische theorieën wordt verwezen.[25] Toch denk ik dat men de rol van de sociobiologie hier niet moet overschatten. Een beetje demagoog slaat net zo veel munt uit het verhaal van onderzoekers die het met hem eens (lijken te) zijn, als uit dat van onderzoekers die hem tegenspreken en die ‘dus’ hun geprivilegieerde positie misbruiken om een onwelkome waarheid te verdoezelen. Bovendien preken partijblaadjes voornamelijk voor eigen parochie. Een groter risico loopt in zekere zin de argeloze lezer van publieksbladen en van wetenschapsrubrieken in de dag- en weekbladpers, waarin op soms half-geamuseerde toon de nieuwste inzichten van sociobiologen worden gepresenteerd. Wie dat regelmatig onder ogen krijgt zou kunnen gaan denken dat die inzichten misschien toch waardevol zijn. En die verleiding wordt, voor wie de nuance zoekt, alleen maar vergroot door de ongenuanceerde scheldpartijen van tegenstanders van de sociobiologie.[26] Het lijkt me daarom zaak zo rustig, afstandelijk en nuchter mogelijk uit te zoeken wat er wel en wat er niet deugt aan sociobiologische verhandelingen.
Toch is daar de kous niet mee af. Stel dat er het een en ander blijkt te schorten aan sociobiologische theorieën, wat zal dan de reactie van voorstanders van dit vak zijn? Het ligt voor de hand dat veel voorstanders net zo zullen reageren als ze ook nu al doen op kritische kanttekeningen: ‘Natuurlijk is de theorie niet volmaakt, er valt nog veel te verbeteren, zo werkt wetenschap immers, steeds opnieuw maken we onze theorieën een beetje beter en langzaam benaderen we de waarheid steeds weer dichter, maar bedenk wel dat de nieuwste theorie ook steeds de beste is die we hebben’. Het motto van veel wetenschappers lijkt soms te zijn: beter de beste theorie die we hebben, al is die misschien verre van volmaakt, dan niets. Een intellectueel horror vacui. Ik kan me in deze visie niet vinden. En dat is dan meteen een reden waarom ik methodologisch onderzoek ook zinnig vind als er geen alternatieven worden geboden. Ik denk dat stilte vaak verre te verkiezen valt boven een niet zo heel erg goede theorie. Wetenschappers zouden veel vaker dan ze nu doen hun mond moeten houden.
1.3. opzet van het onderzoek
Gezien de enorme vloed aan publicaties van en over de sociobiologie is het onmogelijk alles te behandelen. Het was in elk geval mij onmogelijk alles te lezen of zelfs maar vluchtig in te zien en een zorgvuldig overwogen keuze uit het totale aanbod te doen. Ik heb mij beperkt tot een paar, naar ik meen centrale, thema’s en een keuze uit de mij bekende literatuur. Wat dat betreft gaat het hier dus niet over dé sociobiologie maar over de thema’s die ik behandel en hun uitwerking in de literatuur waarnaar ik verwijs.
Hoofdstuk 2 begint met een bespreking van een reeks begrippenparen die in discussies over sociobiologie steeds weer samen opduiken: genetisch bepaald tegenover omgevingsbepaald; biologisch tegenover sociaal; natuurlijk tegenover cultureel; aangeboren tegenover aangeleerd. We zullen zien dat de manier waarop die begrippen gebruikt worden hoofdzakelijk tot verwarring leidt. Na die bespreking sta ik uitgebreid stil bij de relatie tussen ‘cultuur’ en ‘omgeving’. Dat vanwege een groeiend aantal theorieën over culturele evolutie en de evolutie van cultuur. Ik denk dat cultuur in de biologie een overbodig begrip is, en dat dat vak meer gebaat zou zijn met een uitwerking van theorieën over de invloed van de omgeving en van omgevingsveranderingen op de bouw en het gedrag van levende wezens.
In hoofdstuk 3 komt het altruïsmebegrip uitvoerig aan de orde. Ik zal laten zien dat altruïsme in de biologie iets totaal anders betekent dan in de sociale wetenschappen en in het dagelijks leven. Altruïsme kan binnen de biologie op verschillende manieren worden gedefinieerd. Dat leidt tot een reeks verschillende begrippen waarvan sommige zeer abstract en meerplaatsig zijn. Elk van die begrippen is wel in een of andere context bruikbaar, maar men moet niet uit het oog verliezen dat het in feite om verschillende begrippen gaat. Gedrag dat in een ethologische context met recht altruïstisch kan worden genoemd hoeft dat niet ook te zijn in een evolutionair relevante zin des woords.
Verscheidene sociobiologen menen dat uit hun theorieën morele normen kunnen worden afgeleid. En er zijn filosofen die hen hierin volgen. In hoofdstuk 4 bespreek ik een aantal pogingen om de relatie tussen sociobiologie en ethiek uit te werken. Ik zal laten zien dat van een op sociobiologische theorieën gefundeerde ethiek geen sprake is.
Verder menen velen dat inzicht in de menselijke natuur van groot belang is voor morele discussies. En er zijn er die denken dat we dat inzicht aan de sociobiologie kunnen ontlenen. In hoofdstuk 5 zal ik laten zien dat sociobiologie, of ruimer evolutiebiologie, ons weinig over de menselijke natuur kan vertellen. Elke reconstructie van ons evolutionaire verleden zal moeten beginnen bij een (voorwetenschappelijk) beeld van hoe mensen zijn. Dat zal ik aan de hand van een aantal voorbeelden van zulke reconstructies toelichten. Wat die reconstructies ons vervolgens leren over de menselijke natuur verschilt in niets van het mensbeeld waar de auteurs mee begonnen. En daar wordt dat mensbeeld niet wetenschappelijker van. Bovendien zou je je af kunnen vragen of er wel zoiets als dé menselijke natuur bestaat. Ik zal, mede op grond van evolutionaire overwegingen, betogen dat er goede reden is om daaraan te twijfelen.
2 evolutie en cultuur
Heftige discussies over sociobiologie gaan altijd over pogingen om ménselijk gedrag evolutionair te verklaren. Zulke discussies zijn niet nieuw. Darwins ideeën over de mens[27] werden ook al niet overal enthousiast ontvangen.[28] Toch is het niet zo dat elke bioloog die over menselijk gedrag schrijft en een flinke scheut evolutiedenken in zijn verhalen stopt onmiddellijk publiekelijk ter verantwoording wordt geroepen. Desmond Morris’ verhalen[29] leidden tot niet veel meer dan een incidenteel gefronste wenkbrauw, en zelfs de ideeën van Konrad Lorenz,[30] die zich nota bene ooit in geschrifte aanhanger van nazistische rassentheorieën had getoond,[31] riepen niet de storm van protest op die vooral Wilson, maar ook Barash en Dawkins,[32] te verwerken kregen.
Het zou interessant zijn om na te gaan waarom mensen als Lorenz en Morris zo veel minder, en minder zware, kritiek te verwerken kregen dan de volgelingen van Wilson,[33] maar een analyse daarvan ligt niet in mijn bedoeling. Wel wil ik de vinger leggen bij twee duidelijke verschillen. Morris schrijft met liefde over mensen. Hij knaagt wel eens aan onze zelfvoldaanheid, maar dan blijft het bij milde spot. Wilsons toon is een heel andere. Hij ontmaskert liefdeloos. Maar Wilson is wel heel duidelijk over de theorie waarvan hij uitgaat, terwijl Morris zwijgt over theoretische achtergronden. Zonder de ‘genen voor gedrag’ die Wilson expliciet noemt en die tot felle kritiek leidden, zou Morris’ verhaal weinig hout snijden, maar door ze niet te noemen voorkomt de laatste heel wat gekrakeel.
2.1. dichotomieën
Sociobiologen die iets over mensen zeggen worden veelvuldig geconfronteerd met de kritiek dat hun verklaringen gevaarlijk eenzijdig zijn, een bezwaar dat dan vaak als volgt wordt onderbouwd: dierlijk gedrag is aangeboren, dat wil zeggen biologisch of genetisch bepaald, en dus is het prima te verklaren met behulp van biologische theorieën. Het gedrag van mensen daarentegen is aangeleerd en wordt bepaald door hun omgeving, hun cultuur, door sociale factoren, en om die invloeden in rekening te brengen zijn menswetenschappelijke theorieën onontbeerlijk.[34]
Deze kritiek lijkt te berusten op drie vooronderstellingen. Men maakt onderscheid tussen twee typen gedrag: gedrag is aangeboren, natuurlijk, genetisch of biologisch bepaald, óf het is aangeleerd, sociaal, cultureel of door de omgeving bepaald. Vervolgens neemt men aan dat de verschillende begrippen die worden gebruikt om gedrag in één categorie te karakteriseren ongeveer synoniem zijn: aangeboren, genetisch bepaald, biologisch bepaald, natuurlijk, betekenen ongeveer hetzelfde, en dat geldt ook voor aangeleerd, sociaal bepaald, cultureel bepaald, en omgevingsbepaald. Ten slotte gaat men er van uit dat biologen vooral verstand hebben van gedrag uit de eerste categorie, gedrag dat men vooral bij dieren vindt, en beoefenaars van de sociale wetenschappen van gedrag uit de tweede categorie, dat met name kenmerkend voor de mens zou zijn (zie afb. 2.1).

Tegen deze manier van denken wordt regelmatig gewaarschuwd.[35] Auteurs die met deze tweedelingen werken benadrukken veelvuldig dat het niet om absolute tegenstellingen gaat. En zo uitgebreid als ik die tweedelingen hierboven presenteerde vind je ze nergens expliciet.[36] Maar toch blijft het denken in parallelle dichotomieën discussies over sociobiologie beheersen.[37]
Op zich is er niets tegen tweedelingen. Ze verschaffen vaak op een eenvoudige manier een helder overzicht. Maar om ze probleemloos te kunnen gebruiken moeten ze aan twee criteria voldoen. En die criteria gelden evenzeer voor classificaties met méér dan twee categorieën. Een classificatie moet uitputtend zijn, dat wil zeggen dat alle zaken die voorkomen in het domein waarop de indeling betrekking heeft in één der categorieën vallen. De levende natuur werd ooit in planten en dieren verdeeld. Maar de ontdekking van allerlei microscopisch grut leidde tot grote verwarring. Waar hoorden al die eencelligen bij? Pas toen het aantal categorieën werd uitgebreid tot drie (planten, dieren en eencelligen) of zelfs vijf (planten, dieren, eencelligen, prokarioten en virussen) paste alles er weer in.
Verder is het handig als de categorieën in een indeling elkaar uitsluiten. Als je onderscheid maakt tussen mensen en dieren, maar je definieert dieren zodanig dat ook mensen onder die omschrijving vallen, dan kun je onverkwikkelijke ruzies krijgen over de vraag of mensen nu wel of geen dieren zijn. Een complete classificatie met nette, elkaar uitsluitende categorieën voorkomt dat soort onenigheid.
Ik zal nu eerst de dichotomieën van afbeelding 2.1 een voor een bespreken en laten zien dat ze geen van alle veel met de tegenstelling tussen mensen en andere dieren te maken hebben, dus dat ze geen van alle reden geven om de mens apart te zetten als het om evolutionaire verklaringen van gedrag gaat.[38] Daarna zal ik iets meer zeggen over de relatie tussen culturele invloeden en omgevingsinvloeden binnen het kader van (evolutie)biologische theorievorming.
2.1.1. aangeboren en aangeleerd gedrag
De term ‘aangeboren’ verwijst naar alle kenmerken die een individu bij zijn geboorte heeft, ongeacht welke factoren tot die kenmerken hebben geleid. Voor kenmerken die pas later ontstaan gebruikt men vaak de term ‘aangeleerd’. Maar leren is niet de enige factor die een rol speelt bij veranderingen na de geboorte. Secundaire geslachtskenmerken ontwikkelen zich tijdens de puberteit onder invloed van hormonen, daar komt zo direct geen leren aan te pas. En viervingerigheid ten gevolge van een ongeluk is ook niet aangeleerd. Het is daarom beter om van aangeboren en na de geboorte verworven eigenschappen te spreken. Dat levert ons tenminste een uitputtende tweedeling op. Alleen, de tegenstelling tussen aangeboren en verworven eigenschappen is nogal willekeurig. Geboren worden is voor sommige organismen een ingrijpende gebeurtenis, maar niet voor alle organismen, en zeker niet voor alle organismen de énige ingrijpende gebeurtenis. Wat zou je bijvoorbeeld bij vlinders aangeboren eigenschapen moeten noemen? De kenmerken van de vlinder vlak na de verpopping? Of ook die van de rups op het moment dat die uit het ei kruipt?
Wellicht wordt het idee dat gedragskenmerken van dieren vooral aangeboren en die van mensen vooral aangeleerd zijn, gedragen door het grote belang dat leren heeft voor het zich ontwikkelende mensenkind, en dan vooral leren van soortgenoten onder omstandigheden die daar speciaal op zijn toegesneden: onderwijs. Maar zo uniek zijn mensen niet. Heel veel dieren verwerven eigenschappen die ze bij hun geboorte nog niet hadden, bij veel dieren gaat het dan om verwerving langs een weg die we bij mensen ‘leren’ noemen,[39] en dat gebeurt niet zelden onder omstandigheden die daar door bijvoorbeeld de ouders speciaal voor lijken te zijn geschapen. We zouden hoogstens kunnen twisten over de vraag of dieren, net als mensen (soms), begrijpen wat ze geleerd hebben en dát ze iets geleerd hebben. Maar dat zou een twist zijn over iets dat we nu onmogelijk te weten kunnen komen. Hoe dan ook, de mens is net als elk ander dier een lappendeken van aangeboren en verworven eigenschappen. Interessante verschillen zijn er in dat opzicht niet.
2.1.2. genen en omgeving
Om na te gaan hoe zinvol het onderscheid tussen genetisch bepaalde en omgevingsbepaalde kenmerken is, en in hoeverre dat iets met verschillen tussen mensen en andere dieren te maken heeft, moeten we eerst onderscheid maken tussen verschillende betekenissen van de begrippen ‘genetisch bepaald’ en ‘omgevingsbepaald’.
In de biologie bestaan twee vakken die elk genetica heten, de moleculaire genetica, die zich bezig houdt met de bouw van DNA en verwante verbindingen, en hun rol bij de ontwikkeling van individuele organismen, en de populatiegenetica, die zich richt op de kenmerkenverdeling in groepen organismen en de veranderingen van die verdeling van generatie op generatie.
In moleculair-genetisch perspectief kunnen we bij ‘genetisch bepaald’ denken aan zoiets als ‘veroorzaakt door een (of meer) stukje(s) DNA’. Het is echter de vraag of kenmerken in die zin ooit genetisch bepaald kunnen zijn. Een al te eenvoudige beschrijving van de manier waarop een organisme tot stand komt kan suggereren dat het daarbij gaat om het uitvoeren van een programma dat in de vorm van een DNA-streng in de celkern ligt opgeslagen. Stukjes DNA bepalen de bouw van enzymen (en structuureiwitten) en enzymen regelen de ontwikkeling van het organisme en bepalen zo zijn kenmerken. Uiteraard heeft die DNA-streng een bepaalde omgeving nodig, een bepaalde temperatuur, vochtigheid, zuurgraad, enzovoort. En als die omgeving niet helemaal aan de eisen voldoet kunnen er soms rare dingen gebeuren, maar die vallen buiten het normale patroon.
Bij deze manier van beschrijven wordt de informatie in het DNA soms als ‘blauwdruk’ of ‘cake-recept’[40] opgevat. In de blauwdruk-visie wordt het volgroeide individu als het ware onderdeel voor onderdeel beschreven door het DNA.[41] Bij het cake-receptidee gaat men ervan uit dat het DNA niet het eindproduct maar het productieproces beschrijft.[42] Beide[43] beelden geven een te simpele voorstelling van zaken. Een organisme is het resultaat van een zich ontwikkelend embryo, en elke stap in die ontwikkeling hangt af van de structuur van het embryo op dat moment én de waarde van een groot aantal factoren in de omgeving van dat embryo. En in zekere zin gaat die ontwikkeling na de embryonale fase gewoon door, zij het langzamer. Het DNA bepaalt daarbij de samenstelling van gereedschap en bouwmaterialen.[44] Dus als men bij ‘genetische bepaaldheid’ van kenmerken denkt aan de rol van het DNA bij het totstandkomen van een individu dan bestaan er geen genetisch bepaalde kenmerken, en al evenmin omgevingsbepaalde kenmerken.
Populatiegenetici houden zich niet bezig met genen en omgevingsfactoren als oorzaken van kenmerken, maar met genetische verschillen en omgevingsverschillen als oorzaak van kenmerksverschillen. Ongelukkigerwijze laat men het woord ‘verschillen’ meestal weg en spreekt men gemakshalve vaak over genetisch bepaalde kenmerken. Maar wat men met die uitdrukking bedoelt snijdt wel degelijk hout. Een paar voorbeelden kunnen dat duidelijk maken.
Volgens de gangbare theorie zijn verschillen in oogkleur tussen mensen in populatiegenetische zin genetisch bepaald. Weliswaar is de oogkleur van een individu het product van heel veel genen en heel veel omgevingsfactoren, maar het verschil in oogkleur tussen twee individuen is het gevolg van een, relatief klein, genetisch verschil. Bij huidskleur ligt dat net iets anders. Soms zijn huidskleurverschillen net als oogkleurverschillen genetisch bepaald, maar ze kunnen ook heel wel omgevingsbepaald zijn. Een beetje zon maakt van een bleekneus een gebronsde verschijning. Het huidskleurverschil tussen de bleekneus en zijn doorgaans even bleke broertje dat net bruinverbrand van een tropische vakantie terugkomt is omgevingsbepaald, terwijl het even grote verschil tussen de bleekneus en zijn buurman van Noordafrikaanse afkomst genetisch bepaald is. Een en hetzelfde organisme kan dus in verschillende vergelijkingen worden betrokken en het hangt van de vergelijking af of een bepaald kenmerk als genetisch bepaald of omgevingsbepaald (of beide ten dele) uit de bus komt. Door niet duidelijk over verschillen te spreken maakt men het vergelijkingsaspect onzichtbaar. En dat kan tot vervelende redeneerfouten leiden.
Hoe verwarrend het kan zijn om over de genetische bepaaldheid van een kenmerk te spreken blijkt bijvoorbeeld uit een aanval van Vroon op Wilson. Volgens Vroon[45] beweert Wilson dat intelligentie bij de mens genetisch bepaald is. Vroon bestrijdt dat onder verwijzing naar ernstige onvolkomenheden in onderzoek dat had moeten aantonen dat intelligentieverschillen tússen mensen voortkomen uit genetische verschillen. Het zou mij geenszins verbazen als Vroon gelijk had, alleen, dat is hier niet ter zake. Voorzover Wilson[46] iets zegt over intelligentie gaat het blijkens de context vooral over intelligentieverschillen tussen mensen en ándere diersoorten.
We kwamen op deze uitweiding over genetische zaken omdat tegenstanders van een sociobiologische benadering van de mens tegenwierpen dat menselijk gedrag niet, of vaak niet, genetisch bepaald is. Zouden ze daarmee doelen op moleculair-genetische bepaaldheid dan hadden ze op een vrij triviale manier gelijk. Niets is in die zin genetisch bepaald. Dat op zich is geen bezwaar tegen een evolutionaire benadering van menselijk gedrag. Maar ook wanneer ze een populatiegenetische interpretatie op het oog hebben doet hun tegenwerping niet ter zake. Stel dat gedragsverschillen tussen individuen bij de mens vaker omgevingsbepaald zijn dan bij andere diersoorten,[47] dan zou dat nog geen bezwaar zijn tegen een biologische benadering van menselijk gedrag. Biologen besteden regelmatig aandacht aan omgevingseffecten, en veel van hun theorieën houden expliciet rekening met die effecten.
Toch is een verwijzing naar de grote omgevingsbepaalde variatie in gedrag van mensen wel van belang voor sociobiologen. Hun verklaringen mogen die variatie niet uitsluiten. Soms lijken ze dat wél te doen, en dat is een ernstige tekortkoming (ik kom daar in 2.2.2 en 2.2.3 op terug). Maar dat verwijt richt zich op bepaalde theorieën van bepaalde sociobiologen, niet op een evolutionaire verklaring van menselijk gedrag op zich. Evolutiebiologie is moeilijker dan sommige sociobiologen denken, maar er is geen enkele reden om te denken dat de mens uit haar domein ontsnapt is.
2.1.3. natuurlijk en cultureel
Natuur en cultuur hebben beide verschillende betekenissen en bijna al die betekenissen zijn vaag. Natuur betekent soms: bijna alles wat buiten is: planten en dieren, bergen, beken en woestijnen. Maar wat die dingen meer gemeen hebben dan dat ze buiten zijn is onduidelijk. In deze betekenis is natuur dus een wat overbodig begrip. Natuur kan ook de keerzijde van het bóvennatuurlijke zijn. In die zin zijn huizen ook natuur, wat ze in de eerste betekenis duidelijk niet waren, hoewel ze, zowat per definitie, buiten zijn. Natuur kan verder betekenen: alles wat niet, of niet bewust, door mensen gemaakt of bewerkt is. Of woestijnen in die zin natuur zijn is onduidelijk, tenzij we dat woordje ‘bewust’ benadrukken. Woestijnen zijn weliswaar voor een deel het gevolg van menselijke activiteiten maar ze zijn zeker niet bewust gecreëerd. Alleen, als we dat bewust té zeer benadrukken zijn sommige steden ook natuur.[48] Ze zijn soms, net als woestijnen, het onbedoelde effect van menselijk handelen. En de zeer bewust en met zorg gekweekte tulpen waar ons land mee geurt zijn duidelijk níet natuur in deze zin.
Een veelgebruikt argument tegen een sociobiologische benadering van menselijk gedrag is dat menselijk gedrag cultureel bepaald en niet zo maar natuurlijk is. Dat argument vooronderstelt dat natuur en cultuur goed te scheiden zijn. En daar liggen problemen.
Natuurlijk kan ook een synoniem zijn van ‘spontaan’. Zo kunnen we onderscheid maken tussen het natuurlijke gedrag van ijsberen en de dwangmatige rondjes die de ijsbeer in Artis steeds en steeds maar weer zwemt en loopt, en tussen de natuurlijke ontplooiing van een boom die de ruimte heeft en de plat verwrongen takkenbos van een boom in een rotsspleet. Aan onnatuurlijkheid (cultuur?) in deze zin hoeft dus helemaal geen mensenhand te pas te komen. Kortom, wie iets natuurlijk noemt heeft nog heel wat uit te leggen voor duidelijk is wat hij bedoelt.
Ook het begrip ‘cultuur’ kan zeer veel verschillende betekenissen hebben.[49] Die zal ik hier niet allemaal bespreken. Maar en aantal aspecten duiken steeds weer op, die zijn dus kennelijk belangrijk. In de eerste plaats wordt cultuur gebruikt als een verzamelnaam voor menselijk gedrag en de producten daarvan: huizen, gereedschappen, kleren, sieraden, boeken, enzovoort. En dat stelt ons meteen voor een probleem. Er is in elk geval één manier waarop we cultuur en natuur zo kunnen definiëren dat ze tegenover elkaar komen te staan, en dat is door alles cultuur te noemen wat door mensen is gemaakt of onder hun invloed is veranderd, en natuur de hele rest. Bij deze interpretatie van de betreffende begrippen is de mens inderdaad cultuurdrager bij uitstek, maar daaruit volgt nog niet dat de mens buiten het domein van de sociobiologie valt. Een definitie is daarvoor niet voldoende. Het zou zo moeten zijn dat de mens zich feitelijk van andere dieren onderscheidt.
Mensen zijn uitzonderlijk handvaardig en productief, maar op zich is dingen maken geen bij uitstek menselijke bezigheid.[50] Wel valt op dat mensengedrag erg variabel is, en dat geldt ook voor de voorwerpen die zij maken. ’s Mensen variabiliteit in gedrag wordt vaak toegeschreven aan de grote verscheidenheid aan omgevingen waarin mensen voorkomen. Dat is een boeiend gegeven, vooral als je bedenkt dat die omgevingsvariabiliteit op zich weer het gevolg kan zijn van menselijke activiteiten. Sommigen stellen zich voor dat de interactie tussen de mens en zijn omgeving de mens als het ware onttrekt aan het biologische evolutieproces waaraan hij ooit onderworpen was. Zij spreken dan van een proces van culturele selectie dat de biologische selectie aanvult of zelfs tegenwerkt. Die gedachte heeft ook bij sommige sociobiologen ingang gevonden. Ik kom daar later in dit hoofdstuk nog op terug. Laten we intussen niet vergeten dat de mens de enige niet is die in een zelfgemaakte omgeving leeft. Termieten en bevers kunnen hun omgeving snel en drastisch veranderen, en ik vraag me af of niet elk levend wezen in zijn omgeving sporen nalaat die op zich weer van invloed zijn op zijn eigen gedrag en dat van z’n soortgenoten.
Hoe dat ook zij, aan de stelling dat menselijk gedrag niet biologisch verklaard kan worden omdat het cultureel, en niet zo maar natuurlijk is, ligt vaak een wat ‘dieper’ cultuurbegrip ten grondslag.[51] Cultuur verwijst dan naar leerprocessen (die we overigens bij allerlei andere dieren ook zien),[52] en naar de betekenissen die mensen aan alles om hen heen geven. Cultuur wordt in dit verband soms zelfs gezien als een zelfstandige eenheid, een autonoom systeem van betekenisrelaties dat niet zo maar gereduceerd kan worden tot de activiteiten van afzonderlijke individuen die leven binnen een cultuur. Denken over cultuur in deze zin is mijn vak niet, maar ik zal proberen te schetsen wat ik me er bij voorstel (voorzover ik me er iets bij voorstel). Wat voor de een een onbenullig vaasje of een middelmatig boek is, kan voor een ander heel belangrijk zijn, bijvoorbeeld omdat het de enige herinnering is aan een verloren familielid of het eerste geschenk van een geliefde. Zoiets geldt niet alleen voor gebruiksvoorwerpen, maar ook voor bijvoorbeeld gewoontes, uitdrukkingen, religies. En verschillen in betekenistoekenning vind je niet alleen tussen individuen, maar ook tussen groepen. Ik neem aan dat juist die gedeelde betekenissen, voortkomend uit een gedeelde ervaring, voor het cultuurbegrip waar het hier om gaat van belang zijn.
Helaas zie ik niet goed hoe je in dit verband over een cultuur-natuur onderscheid zou moeten denken. In de eerste plaats is er een praktisch probleem. Aanhangers van de opvatting dat cultuur als een betekenissysteem moet worden gezien lijken er soms van uit te gaan dat zoiets bij dieren — dus in de natuur (!?) — niet voorkomt: dieren zouden wel signalen gebruiken, maar symbolen — dingen waarvan de betekenis losstaat van hun aard — komen daar volgens hen niet voor. Daar ga ik niet zo maar in mee. Er zijn dieren, honden bijvoorbeeld, die hun territorium markeren met geurmerken. Je zou die als signalen — dingen met een duidelijke inherénte betekenis — kunnen opvatten. Maar of dat helder is betwijfel ik. Verschillende dieren reageren verschillend op zo’n geurmerk. Een naburige territoriumbezitter stapt er niet licht overheen, zeker niet als ie eerder ruzie met de buurman heeft gehad. Een passerende kat zal eventjes zeer op haar hoede zijn. Het vrouwtje van de territoriumhouder zal er wellicht eerder rustiger van worden. En de territoriumhouder gedraagt zich binnen zijn eigen grenzen met meer bravoure dan daarbuiten. Die reacties hebben alles te maken met vorige ervaringen van de waarnemer, ze zijn dus net zo ‘arbitrair’ als mijn reactie op het vaasje waar ik het eerder over had. Maar misschien is dat niet genoeg om dieren tot cultuurdragers te maken in de ogen van wie bij cultuur aan betekenissystemen denkt. Misschien is een betekenis méér dan de oorzaak van een persoonlijke reactie.
Nu weet ik van mijzelf wat er door mij heen gaat als ik iets in handen neem dat me aan een verloren geliefde herinnert, en ik vermoed dat dat u soms ook zo vergaat. Maar wat de kat denkt die even miauwt bij de lege mand waar haar inmiddels aan de overburen geschonken jonkie lag te slapen, weten we niet. Sommige mensen zullen denken dat er in zo’n kat niet meer omgaat dan hormonen en een nu doelloos moederinstinct. Ze miauwt omdat een bepaalde geur een kettingreactie van zenuwimpulsen teweeg bracht. Mensen die dat zeggen, zeggen meer dan ze kunnen weten. We weten niet of dieren betekenis toekennen aan wat hen omringt, althans we weten niet of ze dat doen op de manier waarop we (elk voor zich) van onszelf weten dat we dat doen. We weten alleen dat ook dieren verschillend reageren op dezelfde dingen, en we kunnen vermoeden dat dat verschil in reactie samenhangt met persoonlijke geschiedenissen. Van wat er echt in dieren omgaat weten we erg weinig. Dus als cultuur meer moet zijn dan een kwestie van gedrag, weten we niet of het bij andere dieren dan de mens voorkomt.
Naast die onzekerheid is er ook een conceptueel probleem. Ik stel me bij betekenis geven met enige moeite wel iets voor, maar wat is het alternatief? Wat zou ‘niet betekenis geven’ moeten betekenen? Iets onbelangrijk vinden? Iets nemen zoals het is? En wat zou het tegenovergestelde van cultuur dan zijn? Voor het zin kan hebben in termen van dit cultuurbegrip over mensen en dieren te praten moet er nog wel het een en ander verhelderd worden.
2.1.4. biologisch en sociaal
Een volgend bezwaar tegen sociobiologische benaderingen van de mens berust op de tegenstelling tussen biologische en sociale factoren. Ook met dat onderscheid heb ik problemen.
Biologie is een vak. Onder biologie in deze zin vallen: vragen, theorieën, hypothesen, beschrijvingen, manieren van benaderen. Een theorie die niet biologisch is kan van alles zijn: fysisch, theologisch, psychologisch, chemisch, antropologisch, enzovoort. Het verwarrende is dat vaknamen soms ook als namen voor dingen, verschijnselen, worden gebruikt. De regels voor dat gebruik zijn alleen niet zo duidelijk. Spieren zijn bijvoorbeeld heel biologisch in deze zin des woords. Komt dat doordat biologen onderzoek aan spieren doen? Dat mag even zo lijken tot je bedenkt dat ook fysici onderzoek aan spieren doen – spieren hebben interessante mechanische eigenschappen – en dat psychologen heel zinnige dingen over de (spier)kramp in mijn schouders kunnen zeggen. Dat vertegenwoordigers van vak X onderzoek aan verschijnsel Y doen maakt Y dus nog niet tot een X-isch fenomeen. Spieren, en alle andere dingen die men vaak ‘biologisch’ noemt, komen alleen bij levende wezens voor. De meest zinnige interpretatie van ‘biologisch’ is hier dan ook wellicht die van ‘biotisch’, van ‘betrekking hebbend op levende wezens’. Maar dan is menselijk gedrag, hoe sociaal het ook mogen wezen, ook biologisch.
Menselijk gedrag is volgens veel tegenstanders van de humane sociobiologie vooral sociaal bepaald,[53] en dus niet biologisch. Zouden ze met ‘biologisch’ hier ‘biotisch’ bedoelen, dan spraken zij wartaal. Zo zal het dus wel niet liggen. En er is een andere interpretatie mogelijk. Wellicht gaat het hen niet om een tegenstelling maar om een niveauverschil. Uitgangspunt is dan een hiërarchie van niveaus: … - fysisch - chemisch - biotisch - (psychisch) - sociaal[54] - … Op elk van die niveaus doen zich volgens sommigen verschijnselen voor die niet in termen van processen op onderliggende niveaus verklaard kunnen worden. Technischer gezegd: (processen op) hogere niveaus kunnen niet tot (processen op) lagere niveaus worden gereduceerd. Ik waag mij niet aan een bespreking van de problemen rond deze visie. Dat is hier ook niet nodig. Stel dat sociale interacties niet verklaard kunnen worden in termen van (eigenschappen van) individuele organismen, en dat daar een apart vak, met eigen theorieën, voor nodig is. Hebben we dan een reden om onderscheid te maken tussen mensen en dieren? Ik denk het niet. Zowel de mens als allerlei andere dieren vertonen sociaal gedrag. Er is geen enkele reden om te veronderstellen dat een theorie over sociaal gedrag niet op elk van die soorten van toepassing zal zijn. Je kunt bekvechten over de vraag of de sociobiologie zo’n theorie biedt. Uit de rest van mijn verhaal zal blijken dat ik daar zo mijn twijfels over heb. En je zou natuurlijk ook nog kunnen twisten over de vraag of de sociobiologie, als ze zo’n theorie biedt, nog wel biologie mag heten. Maar dat wordt een ruzie over woorden. Hoe dan ook, de vaak gebruikte tegenstelling tussen biologische en sociale fenomenen is bij dit alles volstrekt niet verhelderend.
2.1.5. nogmaals dichotomieën
De dichotomieën die de revue passeerden rammelen, en bovendien hebben ze weinig met elkaar te maken. Aangeboren kenmerken zijn moleculair-genetisch gezien niet meer of minder genetisch bepaald dan later verworven eigenschappen, en populatiegenetisch gezien kunnen ze zowel volledig genetisch bepaald als volledig omgevingsbepaald zijn, en alles daartussenin, afhankelijk van de vergelijkingen die we maken. Genetisch bepaalde gedragingen zijn in geen enkel opzicht ‘biologischer’ dan omgevingsbepaalde. Er is weinig reden om aan te nemen dat dieren in het algemeen relatief meer aangeboren eigenschappen hebben dan mensen. En diergedrag is alleen natuurlijker dan mensengedrag als we natuur definitorisch koppelen aan het niet-menselijke. Kortom, er is weinig verband tussen de begrippen die in afb. 2.1 aan de linkerzijde staan. En voor de rechterzijde geldt hetzelfde.
Het opmerkelijke is nu dat niet alleen veel tegenstanders van een (socio)biologische benadering van de mens zich op deze tegenstellingenstapel hebben beroepen, maar veel voorstanders tot voor kort eveneens. Zij beklemtoonden juist hoezeer menselijk gedrag genetisch bepaald is en hoe weinig invloed cultuur wel heeft, om te concluderen dat de mens dier onder de dieren is en dus vooral door biologen bestudeerd moet worden.[55] Maar het tij lijkt iets te keren. Sociobiologen hebben de cultuur ontdekt.
2.2. culturele evolutie
Zoals gezegd, betoogden sociobiologen lange tijd dat cultuur weliswaar bestaat maar toch uiteindelijk niet al te veel invloed heeft. Maar de laatste tijd lijken sommigen hunner de cultuur te hebben herontdekt. In plaats van haar rol te bagatelliseren maken ze theorieën die moeten laten zien dat zij heel wel binnen een biologisch raamwerk past, en ook van pas komt.[56]
Sociobiologen onderkennen meer en meer dat menselijk gedrag, en ook dat van sommige andere dieren, zo divers en veranderlijk is dat gedragsverschillen niet op basis van louter genetische verschillen te verklaren zijn. Ze leiden daaruit af dat het theoretisch kader van de biologie een uitbreiding met ‘cultuur’ behoeft. Ik denk niet dat dat een juiste gevolgtrekking is. Het theoretisch kader van de biologie biedt meer ruimte voor gedragsvariatie dan sommige vooraanstaande sociobiologen beseffen. Ze hebben zich blind gestaard op het belang van genetische factoren, en daarbij zagen ze de rol van omgevingsfactoren over het hoofd. Uit het vervolg zal blijken dat ze dat nóg doen.
Ik zal nu eerst de rol schetsen die gangbare evolutiebiologie aan omgevingsinvloeden toekent. Daarna zal ik enkele voorbeelden van sociobiologische theorieën over culturele evolutie bespreken. Tenslotte komt de algemenere vraag aan bod wat nu precies de relatie is tussen ‘omgeving’ en ‘cultuur’.
2.2.1. omgevingsinvloeden en evolutiebiologie
Omgeving is een relatief begrip. Populatiegenetici die het over omgeving hebben bedoelen daarmee meestal de omgeving van de populaties die ze bestuderen. Embryologen zullen bij omgeving eerder denken aan de omgeving van het zich ontwikkelende individu. En moleculair-biologen denken bij omgeving waarschijnlijk vooral aan de omgeving van de verbindingen waaraan ze onderzoek doen. Al die biologen zullen menen dat omgeving heel belangrijk is, maar ze zullen daar allemaal net iets anders mee bedoelen.
Een individu heeft in de klassieke biologische terminologie een genotype (al zijn genen), een fenotype (alle kenmerken) en een omgeving. Dat is een vrij nette indeling. Alles past erin. En de categorieën grenzen keurig aan elkaar. Met de uitspraak ‘het fenotype is product van genotype en omgeving’ lijkt dus niets aan de hand. Er blijft niets in ongenoemd. Toch kan ze tot misverstand leiden. De omgeving die samen met het genetische materiaal tot een fenotype leidt is niet alleen de omgeving van het individu (dat wat zich buiten de huid van dat individu bevindt — het ‘uitwendige milieu’), maar ook de omgeving van het genetische materiaal (binnen de huid van het individu — het ‘inwendige milieu’). Dus wie bij omgeving aan de omgeving van het individu denkt, laat met de uitspraak ‘het fenotype is product van genotype en omgeving’ heel wat weg, maar dat valt niet direct op.[57] Evolutiebiologen zijn het er zonder uitzondering over eens dat omgeving belangrijk is, en het meest formeel uitgewerkte deel van hun theorie, de populatiegenetica, gaat daar ook steeds over. Maar daarbij gaat het om omgeving in een beperkte zin, en die krijgt dan ook nog een beperkte rol.
Volgens Sober[58] zou een volledige populatiegenetica uit vier theorieën (samenhangende verzamelingen van wetten) moeten bestaan. De eerste theorie (ruwweg de ontwikkelingsbiologie) beschrijft hoe een bevruchte eicel met een bepaald genotype zich in een bepaalde omgeving ontwikkelt tot een compleet individu met een eigen fenotype. Populaties, groepen van zulke individuen, veranderen van samenstelling door verschillen in overlevingskansen van individuen, door migratie, en door andere evolutionaire krachten. De tweede theorie beschrijft hoe dat in z’n werk gaat. De volgende stap in de populatiegenetische cyclus is de aanmaak van een nieuwe generatie. Individuen produceren geslachtscellen, die vervolgens paarsgewijze worden gecombineerd. Zo ontstaan de bevruchte eicellen waarmee de nieuwe generatie begint. De samenstelling van de geslachtscellenpoel wordt bepaald door eigenschappen van de ouders, bijvoorbeeld hun verschil in vruchtbaarheid. De derde theorie beschrijft hoe dat in zijn werk gaat. En de vierde theorie beschrijft de vorming van nieuwe bevruchte eicellen. Bijgaand schema (afb. 2.2) vat het proces dat door al deze theorieën wordt beschreven nog eens kort samen.
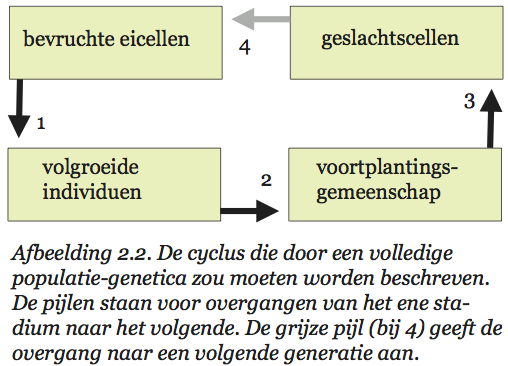
Niet alle door Sober genoemde theorieën zijn even ver ontwikkeld. Aan de eerste theorie, de ontwikkelings-biologie, wordt al heel lang gewerkt. Over de ontwikkeling van embryo’s weten we wel wat, en ook over wat na de embryonale fase gebeurt, is het nodige bekend. Minder uitgewerkt zijn de resterende drie theorieën. In gangbare populatiegenetische modellen worden een aantal van de stappen in het populatiegenetische proces onderdrukt (zie afb. 2.3). Dat kan op twee verschillende manieren zijn uitgewerkt. Soms stelt men een populatie voor als een genotypenverzameling die van generatie op generatie van samenstelling verandert. Sobers vier theorieën worden dan samengevat tot enkele hanteerbare regels die beschrijven hoe uit genotypen door reductiedeling en versmelting van genpakketjes nieuwe genotypen ontstaan, en welke genotypen met welk succes deel hebben aan het ‘voortplantings’-proces.[59] Het alternatief is een beschrijving in louter fenotypische termen. Men modelleert dan hoe opeenvolgende generaties fenotypisch zijn samengesteld, uitgaande van een bepaald (doorgaans genetisch) mechanisme van kenmerkenoverdracht.[60]
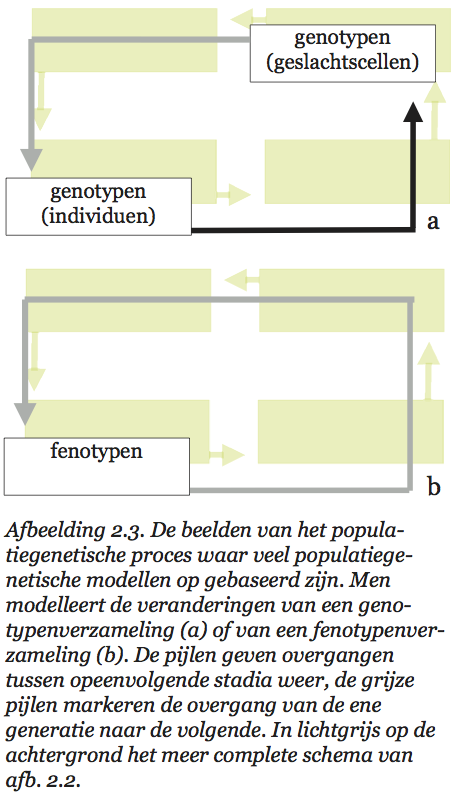
Bij het eerste type modellen (afb. 2.3a) vat men het populatiegenetische proces eigenlijk op als een proces van verandering van de genenpoel van een populatie. De fenotypische veranderin=gen die ons in de eerste plaats aan het denken en onderzoeken zetten beschouwt men dan als de oppervlakkige afspiegeling van een genetisch proces. Het genetische materiaal wordt bij deze vereenvoudigingen de ruggengraat van het leven. Genen zijn de dragers van de informatie die van generatie op generatie wordt doorgegeven, die veranderen kan door mutatie en selectie, en waar alles om draait. En aan het tweede type modellen (afb. 2.3b) ligt in feite vaak hetzelfde beeld ten grondslag. Men modelleert weliswaar veranderingen van fenotypen-verzamelingen, maar meestal gaat men ervan uit dat de verschillen tussen fenotypen uitsluitend genetisch bepaald zijn.
In gangbare populatiegenetische modellen zijn mutatie, selectie, en eventueel drift[61] en migratie — factoren die invloed hebben op de genetische samenstelling van een populatie — de drijvende krachten van het proces. In die factoren is de omgeving in een heel beperkte rol (of scala van rollen) gerepresenteerd. Andere omgevingseffecten blijven buiten beschouwing. Veranderingen in uiterlijk en gedrag die het directe gevolg zijn van omgevingsveranderingen raken uit het zicht. En dat is jammer want zulke veranderingen kunnen heel interessant zijn. En heel algemeen. En ze zouden heel goed consequenties kunnen hebben voor evolutieprocessen. Planten kunnen onder invloed van klimaatsveranderingen vrij drastisch van groeivorm en levenswijze veranderen. Voor dieren geldt dat ook. De aard en de beschikbaarheid van voedsel, het klimaat, het landschap, hebben invloed op grootte, bouw, gedrag, levensloop, voortplantingssnelheid, enzovoort. Ik twijfel er niet aan dat genetische veranderingen een belangrijke rol spelen in het evolutieproces, maar de rol van omgevingsaspecten die direct invloed hebben op het verloop van het individuele ontwikkelingsproces blijven nog te veel buiten beschouwing. De omgeving speelt in populatiegenetische modellen eigenlijk alleen een rol als oorzaak van systematische verschillen in voortplantingssucces tussen verschillende genotypen of fenotypen binnen een populatie, als bron van selectiedruk dus.[62]
Zelfs op Sobers beschrijving van een complete populatiegenetica valt wel wat aan te merken. Hij lijkt zich niet voldoende te realiseren dat niet alleen het genetische materiaal maar ook het ‘interne milieu’ van geslachtscellen een belangrijke rol kan spelen in evolutieprocessen. Ook het interne milieu is een erfenis van vorige generaties, dus met een theorie die het ontstaan van nieuwe geslachtscellen beschrijft ontstaan problemen, als die zich uitsluitend zou uitspreken over de genetische samenstelling van de geslachtscellenpoel. Toch doet Sobers schets, vergeleken bij de gangbare populatiegenetica, en zeker vergeleken bij het deel daarvan waardoor veel sociobiologen zich laten inspireren, de rol van het milieu nog redelijk recht.
Nu is het goed om te bedenken dat populatiegenetici voor een ontzagwekkende taak staan. We zouden te veel eisen als we verlangden dat ze van meet af aan met alle relevante factoren rekening hielden. Het is lastig genoeg om greep te krijgen op de effecten van selectie (verschillen in overlevingskans en vruchtbaarheid). En wie dan ook nog het effect van migratie en van ‘genetic drift’ in z’n beschouwingen wil betrekken, heeft z’n handen echt flink vol. Het is dus niet verwonderlijk dat populatiegenetici omgevingsinvloeden voor een deel negeren of op z’n minst constant veronderstellen. Maar we moeten niet vergeten dat omgevingsinvloeden bij elke stap in het populatiegenetische proces van doorslaggevende betekenis zijn. Ze verdienen een expliciete plaats in ons schema (zie afb. 2.4).
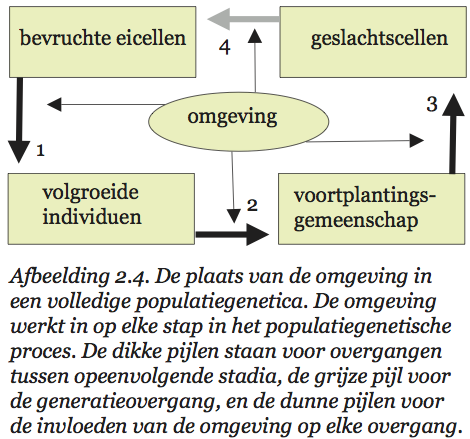
Ik denk dat in dit schema ook alle effecten van ‘culturele’ variatie (althans voorzover de biologie daar iets over zeggen kan) een plaats hebben. Maar laten we eerst eens zien wat het verband is tussen sociobiologische theorieën over culturele evolutie en dit geïdealiseerde beeld van de populatiegenetica.
2.2.2. twee theorieën over culturele evolutie
Ik zal hier twee sociobiologische theorieën over culturele evolutie bespreken, die van Lumsden en Wilson en die van Cavalli-Sforza en Feldman. (Ik doe dat kort, voor een uitgebreider beeld van hun benaderingen, en kritische kanttekeningen daarbij, verwijs ik naar bijlagen 1 en 2, en de daarin geciteerde literatuur.) Die keuze is niet helemaal willekeurig. Het gaat om twee behoorlijk ver uitgewerkte theorieën die onderling voldoende verschillen om een beeld te geven van de diversiteit in deze uithoek van de biologie.
Lumsden en Wilson vatten cultuur op als een verzameling zaken en gedragingen waaruit een organisme kiezen kan. Het gaat daarbij om een grote verscheidenheid aan dingen en handelingen: van technieken om voedsel te vinden en schoon te maken (en daarin vinden we bij veel diersoorten variatie), via manieren om een stropdas te knopen, tot religieuze stromingen. Al die varianten en vormen heten in de terminologie van Lumsden en Wilson cultuurgenen (‘culturgens’). Elk lid van een culturele diersoort leeft in een poel van cultuurgenen. De samenstelling van die poel is kenmerkend voor de betreffende cultuur. Wat een individu doet, hangt af van de cultuurgenen waaruit het kiezen kan en de voorkeuren waarmee het is behept. En door keuzes te maken beïnvloedt het op zijn beurt weer de samenstelling van de hem — en zijn soortgenoten — omringende cultuurgenenpoel. De manier waarop die keuzes tot stand komen vormt dan ook een belangrijk onderdeel van de basis van hun theorie.
Volgens Lumsden en Wilson wordt de keuze van een individu voor een bepaald cultuurgen bepaald door een genetisch vastgelegde voorkeur en de samenstelling van de cultuurgenenpoel. Die voorkeur ligt in de hersenen vast in de vorm van een voorkeursverdeling. Een individu dat geconfronteerd wordt met een verzameling alternatieven heeft voor elk daarvan een bepaalde kans dat hij zich juist dat alternatief zal eigen maken. Doorgaans zal die kans niet 100% voor één alternatief en 0% voor alle andere zijn, hij zal voor meer dan één alternatief groter zijn dan nul. De samenstelling van de genenpoel bepaalt welke alternatieven het individu in welke combinaties zal tegenkomen. Uit die samenstelling, en de voorkeursverdeling van het individu, kunnen we afleiden met welke culturele varianten dat individu door het leven zal gaan. Bijgaand schema (afb. 2.5) laat zien hoe het populatiegenetische model van Lumsden en Wilson er uitziet.
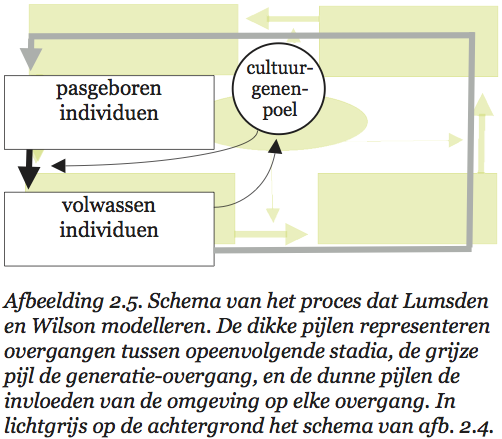
Er zitten een paar problemen aan de aanpak van Lumsden en Wilson. Twee daarvan verdienen hier onze aandacht. De auteurs gaan ervan uit dat pasgeboren individuen over genetisch bepaalde voorkeuren beschikken. We zagen al (in 2.1.2) dat dat een ongelukkige formulering is. Waarschijnlijk zijn de auteurs van mening dat voorkeursverschillen tussen pasgeborenen vaak volledig genetisch bepaald zijn (zie bijlage 1). En dat zou een zeer sterke bewering zijn waarvoor ze geen enkel argument geven. Het lijkt er dus op dat ze de invloed van de omgeving op de ontwikkeling van de bevruchte eicel negeren. De enige omgevingsinvloeden die ze in rekening brengen zijn de invloeden van de cultuurgenenpoel op het volwassen wordende individu. Dat is natuurlijk een aardige aanvulling, maar er is ook hier enige reden voor terughoudendheid. Lumsden en Wilson menen dat die invloeden alleen van belang zijn voor culturele diersoorten, dus voor de mens en enkele andere hogere gewervelden bij wie je iets vindt dat je een culturele traditie kunt noemen. Voor alle andere organismen achten ze de klassieke populatiegenetische benadering voldoende.
Ook voor Cavalli-Sforza en Feldman (zie bijlage 2) is cultuur een keuzekwestie. Na zijn geboorte wordt elk lid van een ‘culturele’ diersoort geconfronteerd met soortgenoten, eerst en vooral de ouders, die zich op verschillende manieren gedragen. Uit dat scala van gedragsmogelijkheden kiest het — bewust of onbewust — een gedragspatroon en maakt zich dat eigen. Die keuze noemen de auteurs culturele selectie en het hele proces van bewustwording, keuze en verwerving, waarin leren een grote rol kan spelen, noemen zij culturele overdracht. Na culturele selectie volgt een proces van natuurlijke selectie: het fenotype van het individu bepaalt zijn overlevings- en voortplantingskans.
Het opmerkelijke is nu dat Cavalli-Sforza en Feldman in hun modellen géén onderscheid maken tussen genetische en culturele overdacht. In die modellen gaat het simpelweg over de fenotypische samenstelling van een populatie en de verandering daarvan in de loop van opeenvolgende generaties. Ze modelleren de invloed van de ouderpopulatie en andere soortgenoten op de ontwikkeling van pasgeborenen (zie afb. 2.6), maar niets in hun modellen suggereert dat het hier uitsluitend om culturele invloeden zou gaan.
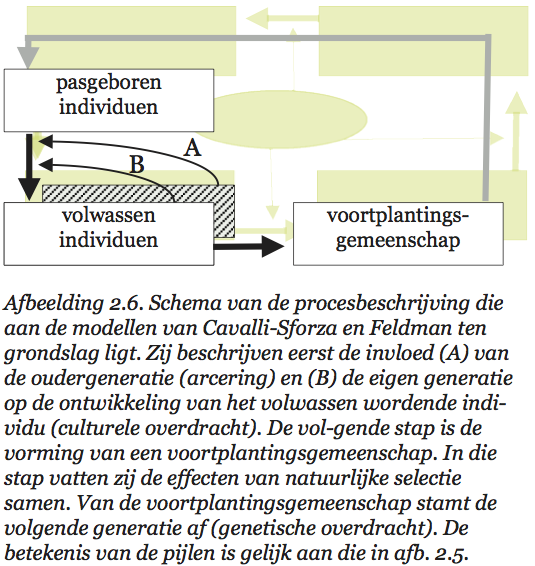
Weliswaar zijn alle voorbeelden die Cavalli-Sforza en Feldman gebruiken ontleend aan wat je ‘cultuur’ zou kunnen noemen, maar hun modellen zijn net zo bruikbaar voor invloeden die niets met cultuur te maken hebben. En daarvan lijken zij zich niet bewust. Zo betogen ze uitgebreid hoe culturele invloeden kunnen leiden tot kenmerken die de kans op nageslacht van het individu bedreigen — het gebruik van alcohol, tabak en ongezond eten, bijvoorbeeld — zonder dat die kenmerken door natuurlijke selectie verdwijnen. Dat is voor hen een belangrijk argument om culturele overdracht als apart mechanisme aan het biologisch arsenaal toe te voegen. Maar ze zien niet dat ook invloeden die hoegenaamd niets met cultuur te maken hebben, besmettelijke ziektes bijvoorbeeld, en die komen bij alle organismen voor, tot ‘fitnessverlagingen’ kunnen leiden. En die verdwijnen ook niet zomaar. In die zin zien ook Cavalli-Sforza en Feldman allerlei omgevingsinvloeden over het hoofd.
2.2.3. cultuur als omgevingsfactor
Evolutiebiologen die evolutie beschouwden als een effect van genotypische veranderingen, liepen vast toen ze beter keken naar de veranderlijkheid van menselijk gedrag. Gedragsveranderingen kunnen heel drastisch zijn, zonder dat er reden is om genetische veranderingen te veronderstellen. Kennelijk, zo redeneren ze nu, komen we daar niet meer uit met genetische informatieoverdracht alleen. En daarom postuleren ze een tweede net van informatieverspreiding: cultuur.[63]
De interpretatie van cultuur als een systeem van informatieoverdracht is niet nieuw in de biologie.[64] En voor wie eerst en vooral in cultuur is geïnteresseerd kan het zinvol zijn eens wat langer met dit idee te spelen.[65] Maar wie primair geïnteresseerd is in evolutie moet toch eerst wat langer nadenken over de relatie tussen cultuur en ‘andere’ biologische factoren.
De gebruiksvoorwerpen en melodieën die Lumsden en Wilson in cultuurgenen hergroeperen zijn omgevingsfactoren voor het zich ontwikkelende individu (en laten we niet vergeten dat ontwikkeling niet een levensfase is, maar iets dat bij alle organismen het hele leven doorgaat). Ze maken deel uit van het fenotype van groepsgenoten, of ze zijn het gevolg van zulke fenotypen (als we gedrag als onderdeel van het fenotype opvatten). Alle organismen beïnvloeden het denken en doen van soortgenoten, en al die invloeden kunnen langetermijneffecten hebben op de fenotypische samenstelling van populaties, dus al die invloeden zullen uiteindelijk door evolutiebiologen moeten worden bestudeerd. Wat ligt meer voor de hand dan dat biologen een theorie ontwikkelen over de directe en indirecte effecten van de fenotypen van soortgenoten op de ontwikkeling van een individu. Als zo’n theorie een beetje volledig is zal zij gaan over de effecten op mijn gedrag van de boeken die ik gelezen heb, over de effecten die de verzorging en zelfverzorging van de moederpoes heeft op haar jongen, over de effecten die een verandering van de vegetatie ten gevolge van het grazen van vorige generaties heeft op de ontwikkeling van jonge zebra’s, enzovoort. Er is geen enkele reden waarom zo’n theorie onderscheid zou moeten maken tussen dingen die we wel en dingen die we niet cultureel noemen. Vanuit het hier geschetste perspectief is er geen wezenlijk onderscheid tussen denneboompjes die van generatie op generatie kleiner worden doordat ze zelf de bodem waarop ze groeien beïnvloeden, en mensen die op een andere manier schilderen of muziek maken dan hun ouders deden doordat ze op een andere manier zijn opgevoed en die daardoor hun eigen kinderen op een weer iets andere manier zullen opvoeden. En voor beide veranderingsprocessen schieten klassieke populatiegenetische verklaringen tekort.
Boyd en Richerson bespreken de relatie tussen omgevingsvariatie en culturele variatie expliciet, maar hun conclusie is een andere dan de mijne. Volgens hen is er een wezenlijk onderscheid tussen culturele variatie en variatie die het gevolg is van wat zij fenotypische flexibiliteit noemen.[66] Culturele variatie wordt van generatie op generatie overgedragen, variatie ten gevolge van fenotypische flexibiliteit — denk bijvoorbeeld aan het formaat van die kleiner wordende denneboompjes — niet. Dat onderscheid zie ik wel, maar het is zaak te zien wat precies het verschil is.
Laten we twee concrete voorbeelden nemen: een moeder die haar dochter cello leert spelen en een kastanjeboom die de grond waarop ze staat uitput zodat het kleine boompje dat zich tussen haar wortels uit een van haar eigen kastanjes ontwikkelt kleiner wordt dan zijzelf is. In beide gevallen is sprake van een ouderlijk fenotype dat een specifiek effect heeft op de omgeving waarin het nageslacht opgroeit en daarmee op het fenotype van het nageslacht. In het eerste geval is de relevante omgevingsfactor de cellospelende moeder zélf. In het tweede geval is het de bodemgesteldheid, dus een efféct van de ouderboom. In het eerste geval lijkt de dochter op haar moeder, er is dus een stukje fenotype ‘overgedragen’, in het tweede geval is die gelijkenis er juist niet, de dochter kastanjeboom is kleiner dan haar moeder en zal dat ook blijven. Dat zijn duidelijke verschillen, maar waarom het wezenlijke verschillen zouden moeten zijn, ontgaat me. Dat te meer daar er allerlei tussenvormen mogelijk zijn. Ik stel me voor dat er zelfs voornamelijk tussenvormen bestaan. Onderwijs verloopt helemaal niet altijd zo direct als in het geval van kinderen die iets door hun ouders zelf krijgen voorgedaan. Allerlei sporen die vorige generaties hebben nagelaten spelen een rol in de ontwikkeling van nieuwe generaties. Bovendien worden ‘stukjes fenotype’, ook in direct onderwijs, maar zelden precies overgedragen. De jonge celliste uit ons voorbeeld zal allerlei in haar ogen ouderwetse fratsen van haar moeder niet overnemen. Alleen al het feit dat haar moeder door zwijmelende aanbidders om haar romantische spel geroemd wordt kan de dochter ertoe brengen uiterst droog en sober te spelen (zelfs als romantiek haar alles behalve wezensvreemd is). Dat doet al een stuk meer denken aan een door ouders uitgeputte bodem! Ik denk daarom dat het beter is om ‘culturele’ gebruiken niet op te vatten als van generatie op generatie overgedragen stukjes fenotype, maar als de fenotypische effecten van de invloed van soortgenoten op de ontwikkeling van nieuwe individuen, als omgevingeffecten dus.
2.3. besluit
Ik ben dit hoofdstuk begonnen met een nette stapel heldere tweedelingen. Die helderheid bleek schijn. Sommige dichotomieën konden op zich nog wel helder gemaakt worden (aangeboren tegenover verworven eigenschappen, genetisch bepaalde tegenover omgevingsbepaalde verschillen), maar bij andere lukte dat alleen met veel gekunsteldheid (natuur tegenover cultuur), of zelfs helemaal niet (biologisch tegenover sociaal). En voor zover het lukte raakte de verwantschap tussen de verschillende tweedelingen volledig zoek. De nette grote tweedeling van afb. 2.1 is dus een grandioze vergissing. Daarmee is aan veel argumenten tegen een biologische bestudering van menselijk gedrag de grond ontvallen.
Dit alles betekent niet dat humane sociobiologie tot nu toe een vlekkeloze onderneming was. Ook sociobiologen kregen in de gaten dat traditionele modellen te simpel zijn voor het verklaren van de diversiteit en veranderlijkheid van menselijk gedrag. Die modellen gaan te veel uit van genetische variatie alleen. De veelzijdigheid van ’s mensen doen en laten was voor sociobiologen een reden om hun modellen in een zeer speciale richting uit te breiden. Ze voegden cultuur toe als een tweede variatiebron. In de theorieën die ze dusdoende ontwikkelden bleek de rol van genetische factoren echter nog steeds aanzienlijk, en dat richtte onze aandacht op wat er ontbrak: het belang van omgevingsinvloeden wordt in theorieën over genetische en culturele evolutie nog steeds ernstig onderschat.
Natuurlijk kunnen we cultuur in evolutiebiologisch perspectief opvatten als onderdeel van de hele klasse van omgevingsinvloeden. Zo bezien is de toevoeging van cultuur aan het evolutiebiologisch begrippenkader een stap in de goede richting. Maar er zitten ook een aantal bezwaren aan die toevoeging. In de eerste plaats is er het risico van begripsverwarring: cultuur in een biologische zin is niet meer dan de schaduw van cultuur in de sociaalwetenschappelijk betekenis (zie 2.1.3). Een tweede bezwaar is belangrijker: voorzover we cultuur als onderdeel van de klasse van omgevingsinvloeden kunnen opvatten is culturele variatie niet meer dan een onderdeel van omgevingsvariatie. Andere omgevingsinvloeden zijn ook heel belangrijk, maar die laten zich niet modelleren middels ‘cultuurgenen’. Een temperatuursverandering is geen potentiële fenotypische eigenschap die een individu al dan niet kan overnemen. We zullen voor die invloeden dus tóch een heel ander soort theorie moeten ontwikkelen. En voor zo’n theorie zou cultuur wel eens niet meer kunnen zijn dan een bijzonder geval.
Ik denk dat evolutionair onderzoek in de toekomst vooral gericht zal moeten zijn op de invloed die omgevingsvariatie heeft op fenotypische variatie. De benadering van Cavalli-Sforza en Feldman laat zien hoe die invloed zou kunnen worden gemodelleerd. Alleen zal die benadering dan wel moeten worden losgeweekt uit het verwarrende culturele jasje waarin de auteurs verstrikt raakten. En wellicht is het verstandig om de mens niet als eerste onderwerp voor onderzoek in deze richting te nemen. De interactie van genetische factoren en omgevingsfactoren bij het totstandkomen van de morfologie van fruitvliegjes is vooralsnog ingewikkeld genoeg.[67]
3 altruïsme: selectie en selectieniveaus
Dieren kunnen verbazend aardig voor elkaar zijn. Iedereen die wel eens huisdieren heeft gehad en daar goed naar gekeken heeft, weet dat. En voor dieren in het wild geldt hetzelfde. Er zijn bijvoorbeeld allerlei vogels die elkaar helpen met nestbouw en zorg voor het nageslacht, of elkaar waarschuwen als er een roofdier nadert.[68] En alle onderzoekers die gedurende lange tijd met zorg de gedragingen van een kleine groep dieren – wolven, chimpansees, gorilla’s – hebben gevolgd en die ‘hun’ dieren bij name kennen, komen thuis met ontroerende verhalen over wederzijdse zorg en aandacht.[69] Natuurlijk vertellen ze ook verhalen over bloedstollende onaardigheid, er is weinig reden om te romantiseren, maar dieren kunnen heel aardig voor elkaar zijn.
Voor sommige mensen is dat een grote opluchting. Vriendelijkheid behoort kennelijk tot de mogelijkheden en wie streeft naar een zorgzame samenleving hoeft niet al van meet af aan het gevoel te hebben dat hij vecht tegen de bierkaai van ’s mensen ‘natuurlijke’ zelfzuchtigheid. Anderen gaan een stapje verder: als zorg voor medeschepselen bij meer dieren voorkomt kunnen we van de biologie vast nog wel wat leren over de omstandigheden waaronder die zorg tot bloei kan komen, en over de grenzen van ónze mogelijkheden. Zij verwachten wat dat betreft vooral veel inzicht te kunnen ontlenen aan de evolutiebiologie.[70] Het evolutieproces immers heeft ons gemaakt tot wat we zijn en is daardoor in laatste instantie bepalend voor onze aard.
Dit soort ideeën wordt vaak heftig bestreden. Men werpt tegen dat de mens nu eenmaal in relevante opzichten van het dier verschilt. Ik heb in het vorige hoofdstuk uiteen gezet waarom die tegenwerping mij niet overtuigt. Of men wijst op het verwerpelijke karakter van de morele voorschriften die voortkwamen uit eerdere pogingen om ethiek een evolutionaire basis te geven: het waren vooral de ideologen van een onbarmhartig kapitalisme, en van het fascisme, die zich op darwinistische ideeën beriepen. Maar ook dat vind ik weinig overtuigend. Als we de evolutietheorie aanvaarden, en áls uit die theorie zou kunnen worden afgeleid dat we de morele plicht hebben vooral voor onszelf op te komen, of het morele recht de zwakkeren om ons heen te verdrukken, dan hebben we ons daarbij neer te leggen. En wie weet volgen er uit de evolutietheorie wel veel vriendelijker normen dan die welke daar eertijds door evolutionaire ethici aan zijn ontleend. De vraag is dus wat de evolutiebiologie ons echt kan leren over de manier waarop we met elkaar zouden moeten omgaan.[71] ‘Moeten omgaan’ vooronderstelt ‘kunnen omgaan’. Leert evolutiebiologie ons iets over wat we kunnen? Voor een antwoord op die vraag zijn een aantal overwegingen van belang.
Veel biologen veronderstellen dat selectie primair aangrijpt op het niveau van het individu. Selectie houdt volgens hen een strijd in tussen individuen, waarbij ieder vecht voor zijn eigen voortplantingssucces. Daaruit zou je kunnen afleiden dat er – biologisch gezien – alleen van egoïsme sprake kan zijn. Deze gedachtengang suggereert dus dat evolutiebiologie niet te verenigen is met de gedachte dat individuen voor andere individuen zullen zorgen. Zoals later zal blijken, liggen de zaken niet zo simpel. Individuele selectie sluit zorg voor anderen niet uit. En daar komt nog iets bij: de veronderstelling dat selectie met name plaatsvindt op het niveau van het individu is problematisch. Er is heel wat verwarring geweest over de vraag op welk niveau selectie zich – uitsluitend of voornamelijk – afspeelt. En die verwarring bestaat nog steeds. Ook dit punt zal ik verder uitwerken.
Er is nog een andere reden om sceptisch te staan tegenover visies op ‘ethisch’ altruïsme gevoed door evolutiebiologie. Ik zal laten zien dat ‘altruïsme’ in verschillende contexten een zeer verschillende betekenis heeft. Wanneer je ‘altruïsme’ zo definieert dat je op evolutionaire gronden iets kunt zeggen over het vóórkomen daarvan, krijg je een begrip dat weinig te maken heeft met altruïsme in een moreel relevante zin. Iets dergelijks zou kunnen gelden voor andere morele categorieën – eerlijkheid, rechtvaardigheid, respect, of liefde, zorg, mededogen.
Daarmee zijn we er niet. Veel moreel relevante categorieën van gedrag mogen evolutionair niet relevant zijn,[72] maar dat betekent natuurlijk nog niet dat de gedragingen die we moreel kunnen karakteriseren zelf ook irrelevant zijn in evolutionair opzicht. Over het morele karákter van gedrag kan de evolutiebiologie ons weinig vertellen, maar over dat gedrag zélf wel. Het kan voor een ethicus die ziet dat zorg voor kleine kinderen doorgaans als een morele plicht wordt beschouwd verhelderend zijn om te weten dat die zorg precies is wat je op evolutionaire gronden van een soort als de onze zou kunnen verwachten. Het feitelijk gedrag, de ouderzorg, kan zo, vanuit een bepaald perspectief, worden verklaard.
Alleen, een verklaring voor het veelvuldig voorkomen van ouderzorg is niet meer dan een verklaring van feitelijk gedrag. Daarmee is nog niet verklaard waarom we ouderzorg goed vinden. Nu is het denkbaar dat ook die evolutionair verklaard zou kunnen worden. Maar daaruit volgt nog niet dat ouderzorg goed is. Wie die stap maakt, de stap van een verklaarbare mening naar morele wenselijkheid, maakt zich volgens veel filosofen schuldig aan wat zij een ‘naturalistische drogreden’ noemen. Of het inderdaad om een drogreden gaat, en of biologen en filosofen die een verbinding leggen tussen evolutiebiologie en ethiek zich aan die drogreden bezondigen, is onderwerp van discussie. Over die discussie gaat hoofdstuk 4. In dit hoofdstuk gaat het uitsluitend over de vraag hoe aardig organismen volgens de biologie voor elkaar kunnen zijn en wat ‘aardig’ hier precies betekent.
3.1. selectieniveaus
Het beeld dat biologen op theoretische gronden hebben van de manier waarop dieren, en dan vooral soortgenoten, met elkaar omgaan schommelt nogal. In het verleden meenden velen dat de natuur gericht is op het in stand houden van soorten. Vanuit die visie ligt enige zorg van soortgenoot voor soortgenoot zeer voor de hand.
Later kwam men tot de overtuiging dat de natuur zo niet werken kan. Een individu dat zou profiteren van de zorgzaamheid van zijn soortgenoten zonder daar veel tegenover te stellen zou duidelijk in het voordeel zijn en zijn zelfzuchtigheid zou zich in de loop der generaties door de hele groep verspreiden, zelfs als dat uiteindelijk tot de ondergang van de groep zou leiden. Om dat te kunnen voorkomen zou de natuur vooruit moeten kunnen zien, en die mogelijkheid past niet in het natuurwetenschappelijk wereldbeeld.
Maar ook de overtuiging dat elk dier uitsluitend op eigen voordeel uit zal zijn, staat inmiddels weer op losse schroeven. Sommige sociobiologen menen dat groepen van individuen onder bepaalde voorwaarden belangrijke eenheden van selectie kunnen zijn. Anderen verdedigen juist dat het bij natuurlijke selectie niet om het individu maar om het gen draait. En paradoxalerwijze zou ook dat tot samenwerking tussen individuen – namelijk tussen dragers van een en hetzelfde gen – kunnen leiden. Nog weer anderen denken dat selectie wel degelijk een kwestie van individuen is, maar dat het die individuen niet zo zeer om hun eigen voortbestaan als wel om het voortbestaan van hun genen gaat, en dus om het welbevinden van alle individuen met wie ze genen gemeen hebben. Op het theorieënfront gaat het heel levendig toe. Ik zal een aantal verschillende visies iets uitgebreider bespreken.
3.1.1. verwantenselectie
Een eenvoudige kijk op het evolutiegebeuren doet vermoeden dat het meeste gedrag van dieren erop gericht is het eigen nakomelingental te vergroten. Maar we zien heel wat dieren dingen doen die dat vermoeden beschamen. Er zijn steeds weer dieren die in plaats van zelf kinderen te krijgen anderen helpen met de zorg voor hun nageslacht, of die hun leven in de waagschaal stellen om soortgenoten uit benarde situaties te redden. Volgens Hamilton[73] is die hulpvaardigheid in veel gevallen heel begrijpelijk, als we maar beseffen dat veel kinderen krijgen niet altijd de hoogste evolutionaire wijsheid is.
Waar gaat het bij evolutie door natuurlijke selectie precies om? Het klassieke verhaal is vrij eenvoudig. Kenmerken van individuen die veel kinderen krijgen zullen in de volgende generatie algemener zijn dan kenmerken van individuen die weinig kinderen krijgen. Aangenomen tenminste dat ouders en kinderen op elkaar lijken. Laten we ons voor de zekerheid beperken tot kenmerken waarbij de verschillen binnen de populatie genetisch bepaald zijn.[74] Kenmerksvarianten – of de daarbij behorende genetische varianten, dat komt nu op hetzelfde neer – die hun dragers relatief kinderrijk maken zullen steeds algemener worden. En we hebben reden te verwachten dat elk individu er als het ware op gericht zal zijn – ‘genetisch geprogrammeerd zal zijn’ is hier de veel gebruikte maar hoogst ongelukkige term – om zo veel mogelijk nageslacht te verwerven. Het is Hamiltons verdienste dat hij inzag dat er in dit verhaal een kleine denkfout zit. Mijn kinderen zijn niet belangrijk voor mijn genen omdat ze mijn kinderen zijn, maar omdat ze drágers van (een deel van) mijn genen zijn. In dat opzicht zijn ze niet uniek. Met elk van mijn familieleden heb ik een deel van mijn genetisch materiaal gemeen (de grootte van dat deel hangt af van de mate van verwantschap). In omstandigheden waarin zelf kinderen krijgen relatief veel energie kost kan het daarom handiger zijn om je te richten op de zorg voor andere verwanten.
Hamiltons ideeën geven aanleiding tot veel ingewikkeld rekenwerk. Het is zaak om nauwkeurig te schatten hoe groot de kans is dat een kopie van een willekeurig gen van een individu voorkomt in elk van zijn verwanten. Die kans wordt verwantschapscoëfficiënt genoemd. Hij hangt direct af van de mate van verwantschap tussen de betrokken individuen. Ruwweg[75] kun je zeggen dat die kans ten opzichte van een kind een half is, en dat is hij ook ten opzichte van broers en zussen. Ten opzichte van de kinderen van die broers en zussen is hij een kwart. Dat betekent bijvoorbeeld dat wie de energie die het kosten zou om zelf één kind te krijgen en groot te brengen ook kan besteden aan het opvoeden van drie neven en nichten die anders het loodje zouden leggen, beter voor dat laatste kiezen kan. Evolutionair levert dat meer op: stellen we de waarde van een kind op een half dan zijn die drie neefjes en nichtjes op dezelfde schaal samen driekwart waard. Over die schaal nu eerst iets meer.
In de klassieke evolutiebiologie worden de evolutionaire vooruitzichten van een individu uitgedrukt in wat men noemt de ‘fitness’[76] van dat individu. De fitness van een individu is gelijk aan het te verwachten aantal nakomelingen van dat individu. Ik zal straks (zie 3.2.2.1.3) laten zien dat aan het fitnessbegrip zeer veel haken en ogen zitten. Hier is het voldoende om te weten dat biologen het fitnessbegrip in praktijk vaak interpreteren als (een schatting van) het aantal nakomelingen dat een individu uiteindelijk krijgen zal. Uitgaande van die interpretatie is de fitness van een werkbij nul, ’t arme beest is immers steriel, en die van een bijenkoningin gigantisch. Hamiltons overwegingen laten zien dat de fitness van een individu, op deze manier geïnterpreteerd, geen goede maat is voor zijn evolutionaire vooruitzichten. Andere verwanten dan het eigen directe nageslacht blijven buiten beschouwing. Om dat te ondervangen stelde Hamilton een nieuw begrip voor: de inclusieve fitness, ruwweg gedefinieerd als de som van eigen nakomelingen en die van alle andere verwanten, waarbij voor elk type verwant het aantal gecorrigeerd moet worden voor de mate van verwantschap. De nakomelingen van broertjes en zusjes tellen maar voor de helft mee, en die van neefjes en nichtjes voor een kwart, enzovoort. Elk individu zou er volgens Hamilton op uit moeten zijn z’n eigen inclusieve fitness te maximaliseren. Om het verschil tussen deze visie en de gangbare kijk op selectie aan te geven wordt selectie zoals die volgens Hamilton verloopt vaak aangeduid met de term verwantenselectie (‘kin selection’).[77]
De vraag is nu of dieren die voor soortgenoten zorgen inderdaad bezig zijn om hun inclusieve fitness te vergroten. Er zijn een aantal beroemde voorbeelden die daar op wijzen. Vogels die soortgenoten helpen met de bouw van het nest en de zorg voor het nageslacht, helpen vooral nauwe verwanten, hun ouders bijvoorbeeld. Het gaat daarbij meestal om vogels die een eigen territorium nodig hebben om een gezinnetje op te zetten, en de ‘helpers’ zijn meestal nog te jong en te onervaren om zo’n territorium te kunnen bemachtigen en verdedigen. Zelf kinderen krijgen en grootbrengen is voor hen dus nog niet mogelijk.[78] De werkverdeling bij veel sociale insecten laat zich in Hamiltons perspectief eveneens aardig duiden. Steriele werkbijen helpen bij het grootbrengen van broers en zusters waarvan sommige te zijner tijd voor een nieuwe generatie zullen zorgen. De inclusieve fitness van bijenwerksters is dus bepaald niet nul. Integendeel, door de merkwaardige voortplantingsgenetica van deze insecten zijn ze met hun zusters nauwer verwant dan ze met hun eigen kinderen zouden zijn.[79] Hun inclusieve fitness is daardoor des te groter.
Helemaal onweersprekelijk is Hamiltons vernieuwing niet. Het gaat inderdaad om een grote stap voorwaarts, maar alleen ten opzichte van een zeer eenvoudige kijk op het evolutiegebeuren. Wie fitness opvat als het feitelijke, of te verwachten, kindertal van een individu, en denkt dat selectie gepaard gaat met het maximaliseren van de eigen fitness, maakt de denkfout die Hamilton signaleerde. Maar populatiegenetici werken al heel lang met andere maten, de fitness van een genotype of de genotypische fitness van een individu.[80] En met modellen waarin genotypische fitness als maat voor evolutionair succes wordt gehanteerd kunnen alle verschijnselen die we met behulp van inclusieve-fitnessmodellen kunnen verklaren, ook worden verklaard.[81] We zullen later zien dat zelfs ‘individuele’ fitness op een zinnige manier zo gepreciseerd kan worden dat die kan worden gebruikt voor een verklaring van de hulpvaardigheid waar Hamilton zich het hoofd over brak. Het begrip inclusieve fitness heeft een belangrijke rol gespeeld in de populatiegenetica omdat het beperkingen zichtbaar maakte van gangbare opvattingen over fitness in de oorspronkelijke zin des woords. Maar het ziet er naar uit dat het bij een verscherping van de oude theorieën overbodig wordt. Zo bezien was het een nuttige catalisator, en niet meer dan dat.
3.1.2. groepselectie
Is het denkbaar dat wederzijdse hulp van dieren kan blijven bestaan omdat een groep van dieren die rekening met elkaar houden betere evolutionaire vooruitzichten heeft dan groepen waarbinnen ieder alleen aan zichzelf denkt? In het verleden is deze gedachte door verscheidene biologen met verve verdedigd.[82] Er zijn veldgegevens die suggereren dat bij bijvoorbeeld herkauwers, die in grote groepen leven, het geboorteaantal terugloopt nog voor er van voedselschaarste sprake is. Het lijkt alsof ze hun voortplanting ‘vrijwillig’ beperken om het voortbestaan van de kudde niet in gevaar te brengen. Daar valt wel wat tegen in te brengen. Een individu dat zich niets van die dreigende voedselschaarste aantrekt brengt weliswaar de kudde in gevaar maar vergroot tegelijkertijd de kans dat énkele van haar eigen nakomelingen een periode van honger zullen overleven, gewoon omdat zij meer nakomelingen hééft. De brutaalsten krijgen uiteindelijk veel meer dan de halve wereld. Op grond van die overweging wezen veel biologen groepselectie tot voor kort zonder meer af.
Inmiddels wordt over de rol van groepen genuanceerder gedacht. Er is een hele reeks populatiegenetische modellen ontwikkeld die laten zien dat hulp binnen een min of meer gesloten groep evolutionair geen doodlopende weg hoeft te zijn.[83] Het gaat daarbij doorgaans om modellen van populaties die zijn opgedeeld in veel kleine, tijdelijke groepjes. Telkens als een groep door het overlijden van zijn leden verdwijnt ontstaat een lege plek die wordt opgevuld door kolonisatie vanuit een nog bestaande groep. Men gaat ervan uit dat de levensduur van een groepje groter wordt naarmate het meer ‘altruïstische’ individuen bevat. Met de levensduur neemt ook de kans toe dat een groep de leverancier wordt van kolonisten van opengevallen plekken. Nu zal binnen elke groep gelden dat individuen die zich uitsluitend op eigen voordeel richten op de korte termijn in het voordeel zijn ten opzichte van groepsgenoten die dat niet doen. Maar als de eersten de toon gaan aangeven is het met de groep in z’n geheel gedaan. Men spreekt bij dit soort modellen over ‘groepselectie’ omdat de overlevingskans van de groep (mede)bepalend is voor de uitkomst van het proces.
In modellen van groepselectie zitten nogal wat variabelen: de snelheid waarmee egoïsten een groep kunnen overnemen, de aantalsverhouding van egoïsten en altruïsten binnen de populatie, de kans dat een egoïst door mutatie altruïstische nakomelingen krijgt of omgekeerd, de gemiddelde levensduur van verschillende groepen, en vast nog veel meer. Het zal duidelijk zijn dat er met de waarde van al die variabelen heel wat te spelen valt. Er zijn combinaties van waarden waarbij hulpvaardigheid kan blijven bestaan. Maar het is veel te vroeg om vast te stellen in hoeverre groepselectiemodellen goede beschrijvingen van bestaande populaties zijn.
3.1.3. selectie op verschillende niveaus
Volgens Lewontin treedt evolutie door natuurlijke selectie op als aan drie voorwaarden is voldaan:[84] er dient sprake te zijn van fenotypische variatie binnen een groep; de verschillende vormen, fenotypen, dienen te verschillen in levensduur en voortplantingssucces, in fitness dus; en er moet sprake zijn van een zekere overeenkomst in fitness[85] tussen ouders en kinderen. Hoe die overeenkomst tot stand komt is niet zo belangrijk.
Dit soort principes geldt natuurlijk evenzeer voor andere eenheden dan individuen. Ook genen, cellen, populaties of soorten, verschillen onderling in eigenschappen en in levensvatbaarheid, en ook bij zulke eenheden zou je kunnen spreken van erfelijkheid van fitnessverschillen.[86] Lewontin meent dan ook dat selectie in principe op al die niveaus plaats kan vinden. En hij staat daarin niet alleen.[87]
Lewontin presenteert een heel rijtje eenheden: genen, cellen, individuen, groepen, populaties, soorten en oecosystemen. En volgens hem kan op het niveau van elk van die eenheden selectie plaatsvinden, al zal het individu meestal het belangrijkste niveau zijn. Gezien zijn omschrijving van ‘selectie’ is daar ook wellicht niet zo heel veel tegen in te brengen, al is het te vroeg om heel besliste uitspraken te doen over het precieze belang van alle genoemde niveaus. Maar er is één punt dat mij blijft bevreemden: soorten (kunnen) bestaan uit populaties, populaties (kunnen) bestaan uit groepen, en groepen uit individuen en die weer uit cellen. Maar een cel bestaat niet alleen uit genen, en een oecosysteem bestaat uit populaties en hun omgeving, niet uit soorten. Dat lijkt misschien voer voor kniesoren, maar we zullen zo meteen zien dat in elk geval het feit dat cellen niet uit genen bestaan wel degelijk van belang is. Genen horen in het rijtje van genen via individuen tot soorten beslist niet thuis.
3.1.4. het gen als eenheid van selectie
Volgens Richard Dawkins[88] heeft het eigenlijk alleen maar zin om over selectie te spreken in verband met genen. Daarmee neemt hij als evolutietheoreticus een vrij geïsoleerde plaats in.[89] Zijn analyse van het selectieproces begint met een algemene omschrijving van wat hij onder selectie verstaat, en die omschrijving is veel strakker dan die van anderen.
Dawkins definieert ‘selectie’ in termen van ‘replicatoren’. Elk ding waarvan met een bepaalde snelheid precieze kopieën worden gemaakt is een replicator. Stukjes DNA, genen dus, zijn replicatoren, zo ook teksten die gefotokopieerd worden. Selectie treedt op wanneer er van een bepaalde replicator varianten in omloop zijn die onderling verschillen in levensduur of in de snelheid waarmee nieuwe kopieën ontstaan. Dan namelijk zal de samenstelling van de replicator-‘populatie’ in de loop van de tijd veranderen.
Kopieerprecisie is een noodzakelijke eigenschap van een ‘eenheid van selectie’ in Dawkins’ zin des woords. Die noodzaak is gegeven met zijn definitie van selectie. Individuele organismen, populaties of soorten worden nooit precies gekopieerd, ze kunnen dus per definitie geen eenheid van selectie zijn. Ze spelen natuurlijk wel een rol in het proces dat tot selectie leidt. Dawkins noemt ze ‘voertuigen’ van – in dit geval genetische – replicatoren. De aard en de samenstelling van het biotische voertuigenpark om ons heen verandert onder invloed van selectie op genen. Ook Dawkins ziet wel in dat individuen, of soorten, kunnen verschillen in de snelheid waarmee ze zich ‘vermenigvuldigen’ en dat die verschillen er evolutionair gesproken toe doen, maar ze doen er volgens hem slechts toe voorzover ze leiden tot verschillen in kopieersucces tussen de genen waarvan ze het voertuig zijn.
Dawkins’ visie lijkt een beetje op die van Hamilton. Ook bij hem ging het over individuen die zich niet zo zeer zorgen moeten maken over de toekomst van hun eigen nageslacht als wel over de toekomst van dragers van hun genen. Toch is er wel enig verschil. In Hamiltons verhaal gaat het over individuen die verschillen in de mate waarin ze hun inclusieve fitness weten te maximaliseren. En de succesvolsten beërven de wereld. Populair gezegd: ik moet er voor zorgen een zo groot mogelijke inclusieve fitness te krijgen. Bij Dawkins gaat het over genen die verschillen in hun vermogen het aantal exacte kopieën van henzelf in de volgende genengeneratie zo groot mogelijk te krijgen. Dus: elk van mijn genen moet ervoor zorgen mijn gedrag zo te beïnvloeden dat dat tot een zo groot mogelijk aantal kopieën van dat gen leidt. En het daarvoor benodigde gedrag is niet noodzakelijk voor elk van mijn genen hetzelfde. Het gen dat mij en mijn beide zusters lichtelijk bijziende maakt (stel dat er zo’n gen is) heeft er belang bij dat ik me meer inspan voor hun reproductieve succes dan voor dat van mijn vérziende broer. Voor de genen op mijn Y-chromosoom, die ik wel met mijn broer maar zeker niet met mijn zusters gemeen heb, ligt dat beduidend anders.
Kortom, in een Hamiltoniaanse optiek heeft elk individu een duidelijke opdracht in het leven: maximaliseer je inclusieve fitness. In een Dawkinsiaanse optiek ontbreekt die opdracht. Elk individu wordt rondgecommandeerd door een grote groep genen die elkaar soms danig tegenspreken.[90] Voordat valt vast te stellen wie van beide, Hamilton of Dawkins, gelijk heeft, zullen we zowel meer moeten weten over wat er in de wereld gebeurt als over de precieze betekenis van de beweringen die hier in het geding zijn.
Het loont de moeite om Dawkins’ verhaal nog even te vervolgen. Hij speelt op amusante wijze met de mogelijkheid dat ook genen in andere voertuigen invloed hebben op voorkomen en gedrag van een bepaald individu. Een voorbeeld (niet van Dawkins) ter verduidelijking. Voor het gen dat mijn kat er net iets leuker uit doet zien dan andere katten, en dat mij er daardoor toe brengt net iets beter voor haar (de kat) te zorgen dan ik anders gedaan zou hebben, is mijn betere zorg een deel van zijn effecten (de effecten van het gen) op de wereld. In dit geval een effect dat in het voordeel van het betreffende gen is. Een gen in katten dat mij en anderen tot grote zorgzaamheid brengt is in het voordeel ten opzichte van genen op dezelfde plek[91] die dat effect niet hebben. Genen hebben invloeden die verder reiken dan de grenzen van hun eigen voertuig. Dawkins voert daarom een nieuw begrip in, het uitgebreide fenotype (‘extended phenotype’) van een gen, dat hij definieert als de som van alle effecten van dat gen op de wereld om hem heen.[92] En hij stelt dat een kenmerk van een individu niet begrepen moet worden vanuit de belangen van de genen waarvan dat individu het voertuig is, maar vanuit de belangen van alle genen die op dat kenmerk invloed hebben. En die invloed overschrijdt de grenzen tussen voertuigen, zoals mijn kattenvoorbeeld liet zien.
Dawkins maakt duidelijk dat er in het debat over selectieniveaus twee vragen door elkaar spelen. De vraag welke voertuigen evolutionair gezien het meest relevant zijn, individuele organismen, of zelfs onderdelen daarvan, of groepen van organismen. En de vraag of je het, pratend over selectie, over voertuigen of over replicatoren (dingen dus met een heel hoge kopieerbetrouwbaarheid) moet hebben.
Zijn antwoord op de eerste vraag verschilt niet noemenswaardig van Lewontins opvatting over selectieniveaus. Alleen begint zijn lijstje van eenheden niet met het gen maar met de cel, en dat is netter. We kunnen nu ook zien waarom. In Lewontins lijstje staan replicatoren (genen) en voertuigen (alle andere ‘niveaus’) door elkaar, bij Dawkins zijn die keurig gescheiden.
Echt belangrijk wordt het pas bij de tweede vraag, zij het vooral omdat die vraag door anderen helemaal niet gesteld wordt. Maar Dawkins’ antwoord – het gaat bij natuurlijke selectie om replicatoren en niet om voertuigen – is niet helemaal overtuigend. Hij krijgt zijn gelijk, zoals we in het begin van deze paragraaf zagen, vooral door ‘selectie’ te definiëren in termen van ‘replicatoren’. En waar dat precies voor nodig is maakt hij niet duidelijk. Als de kopieerprecisie nul is zullen verschillen in kopieersnelheid tussen twee eenheden uiteraard weinig sporen nalaten, maar dat is geen reden om te verlangen dat hij honderd procent is. Zolang nakomelingen iets meer op hun eigen ouders lijken dan op andere leden van de ouderpopulatie mag je verwachten dat verschillen in kindertal tussen ouders effect hebben op de eigenschappenverdeling in volgende generaties. Er lijkt weinig op tegen om daarbij van selectie te spreken, en niets sluit uit dat over selectie in die zin een boeiende theorie valt op te stellen.[93]
Kortom, Dawkins schept wat helderheid, maar veel dichter bij een antwoord op de vraag welke eenheden nu een hoofdrol moeten spelen in een theorie over organische evolutie en hoe de bijrollen verdeeld zullen zijn, brengt hij ons niet.
Het enige wat ons na dit overzicht rest is een negatieve conclusie, maar dat is voor het vervolg van dit verhaal genoeg. Niemand verdedigt meer de stelling dat de eigenschappen van een individu dat niet uitsluitend op eigen voordeel uit is, dientengevolge zullen worden weggeselecteerd: dieren die aardig voor elkaar zijn, zijn geen evolutionaire onmogelijkheid.[94]
3.2. altruïsme: begripsvorming
Ik heb het tot nu toe veelvuldig gehad over beesten die ‘aardig zijn’ voor elkaar, die elkaar ‘helpen’, maar dat zijn vage en weinig biologische begrippen. Sociobiologen hebben daarom een wat technischer begripsapparaat uitgewerkt. En de vraag is nu of we de conclusie van de vorige paragraaf in sociobiologische termen kunnen vertalen. Voor we die vraag kunnen beantwoorden zullen we dat begripsapparaat nauwkeurig moeten bezien.
De dieren die ik hierboven aardig noemde noemen sociobiologen ‘altruïstisch’. Dit woord is ontleend aan de vaktaal van de menswetenschappen. ‘Altruïsme’ (letterlijk: gerichtheid op de ander) heeft in de sociale wetenschappen en de ethiek een heel specifieke betekenis: een altruïst is iemand die iets doet met de bedoeling, de intentie, iemand anders terwille te zijn. Nu denken biologen niet allemaal hetzelfde over de vraag of het mogelijk is op een betekenisvolle manier over ‘intenties’ en ‘bedoelingen’ te praten.[95] En niet alleen biologen hebben het daar moeilijk mee, ook in de psychologie staat het gebruik van zulke begrippen ter discussie. Een simpele en effectieve manier om niet door zulke problemen te worden geplaagd is ze eenvoudigweg te omzeilen. Om die reden wordt altruïsme in de biologie vaak op een iets andere manier gedefinieerd, en wel als gedrag dat voor de ‘gedrager’ nadelig is en dat een ander voordeel oplevert. Voor- en nadeel worden meestal gemeten in termen van fitnesseffecten.[96]
Tegenover altruïsme in deze betekenis staat ‘egoïsme’, gedrag dat voor de gedrager voordelig en voor een ander nadelig is. Daarnaast onderscheiden sociobiologen soms nog ‘kwaadaardigheid’,[97] gedrag dat voor beide nadelig is, en ‘samenwerking’,[98] gedrag waar beide voordeel van hebben. Als we ervan uitgaan, en dat doen sociobiologen kennelijk, dat gedrag maar heel zelden geen enkel fitnesseffect heeft, dan vormen deze vier begrippen samen zo op het eerste gezicht een nette indeling van gedragstypen. We zullen straks zien dat dat wat tegenvalt.
3.2.1. van altruïsme naar bioaltruïsme
Sommige sociobiologen menen dat hun definitie van altruïsme in termen van de fitnesseffecten van gedrag niet meer is dan een technische uitwerking van het daagse altruïsmebegrip.[99] Maar dat klopt niet helemaal. De bedoeling waarmee je iets doet sluit namelijk lang niet altijd aan bij de effecten van wat je doet.[100] Een simpel voorbeeld kan dat illustreren.
Als u een eindje omloopt om een oude bekende die slecht ter been is wat boodschappen thuis te bezorgen en u laat onderweg de doos waarin u ze vervoert een paar keer vallen zodat uw kennis later bij het uitpakken haar nieuwe serviesgoed aan gruzelementen vindt, dan bent u in de daagse zin des woords wellicht heel altruïstisch geweest. U wou alleen maar helpen. Maar in de sociobiologische zin des woords was wat u deed misschien wel eerder kwaadaardig, u en uw kennis hebben er beiden nadeel van. Aangenomen tenminste dat een extra stukje lopen negatieve fitnessconsequenties heeft en het teloorgaan van net gekocht serviesgoed ook. Maar dat hoeft niet zo te zijn. Stel dat dat geloop juist heel gezond voor u is. Dan zou uw gedrag, in de biologische zin des woords, egoïstisch zijn. En het is zelfs mogelijk dat u uw kennis van een nare trage dood hebt gered omdat het servies met een ondeugdelijk loodhoudend glazuur was afgewerkt. In dat geval zou er sprake kunnen zijn van samenwerking tot wederzijds voordeel.
Dit voorbeeld is natuurlijk erg simpel. Geen sociobioloog zal een individu op grond van één zo’n gebeurtenis meteen een altruïst of een egoïst noemen.[101] Maar een ding is duidelijk: er is geen noodzakelijk verband tussen de bedoelingen waarmee je iets doet en de effecten van wat je doet. Er is dus geen direct verband tussen de daagse betekenis van begrippen als altruïsme en egoïsme, en die van hun sociobiologische naamgenoten. Om verwarring te voorkomen zal ik, waar nodig, het biologische altruïsmebegrip aanduiden met ‘bioaltruïsme’.[102] Over dát begrip zal het hier verder voornamelijk gaan, maar veel van wat ik schrijf geldt ook voor de andere begrippen uit onze vierdeling. Ik vertrouw erop dat u dat verder zelf kunt invullen.
3.2.2. verschillende bioaltruïsmebegrippen
Bioaltruïsme wordt meestal gedefinieerd voor het gedrag en de fitness van een individueel organisme, en de fitness van een ander organisme. Maar biologen gebruiken het, soms ongemerkt, ook in andere betekenissen. Zowel de gedrager, als de beschrijving van wat die doet, en de munteenheid waarin men de rekening opmaakt (hierboven dus de fitness) kunnen door andersoortige zaken worden vervangen.
Allereerst spreken biologen niet alleen over altruïstische individuen maar ook over altruïstische genen, genotypen of fenotypen (zie 3.2.2.1.1, ik ga er verder stilzwijgend van uit dat de andere partij, degene die profiteert van het altruïsme, dan ook een gen, genotype of fenotype zal zijn). Verder heeft gedrag twee verschillende betekenissen: de manier van doen van een organisme in het algemeen, en een specifieke gedraging op een bepaald moment[103] (zie 3.2.2.1.2). En tenslotte worden naast fitness ook vruchtbaarheid, overlevingskans, of welstand, al dan niet in overdrachtelijke zin, als maat gebruikt,[104] bovendien kan ‘fitness’ op verschillende, elk op zich relevante, manieren worden gedefinieerd (zie 3.2.2.1.3).
Het zal duidelijk zijn dat met behulp van deze verzameling variabelen een lange reeks van verschillende altruïsmebegrippen kan worden opgesteld. Sommige van die begrippen zijn evident onzinnig: niemand stelt zich iets voor bij een individuele gedraging van een genotype en de gevolgen daarvan voor zijn welstand. Voor ik straks wat nettere definities ga uitwerken kunnen er heel wat combinaties worden weggewied. We zullen dan vijf altruïsmebegrippen overhouden die elk op zich bruikbaar zijn maar die wel in belangrijke opzichten van elkaar verschillen (zie 3.2.2.2). Over de implicaties van die verschillen later meer (zie 3.2.3).
Ik zal nu eerst de verschillende variabelen in het raamwerk voor bioaltruïsmedefinities één voor één wat uitgebreider aan u voorstellen.
3.2.2.1.1. hoofdrolspelers in een altruïsmedefinitie
We zagen al dat individuele organismen altruïstisch kunnen worden genoemd. Maar daarnaast treden bij sommige auteurs soms ook genotypen, fenotypen of zelfs losse genen als altruïsten op. Wat zijn dat voor dingen?
Als biologen verklaringen geven van kenmerksveranderingen binnen een populatie hebben ze het nooit over alle kenmerken van de leden van de populatie tegelijk. Heel vaak gaat het slechts over één kenmerk. Ze kijken bijvoorbeeld naar de oogkleur van de cactusmus,[105] en vragen zich af waarom er bij deze vogels verschillende oogkleurtypen voorkomen terwijl één daarvan toch zo duidelijk de voorkeur verdient.
Cactusmussen hebben grijze, groene, of rode ogen, en het verschil tussen individuen met verschillende oogkleur wordt bepaald door twee genen op één locus.[106] Er zijn drie genencombinaties, OO, Oo en oo. Individuen met OO hebben grijze ogen, die met Oo groene, en met oo rode. Roodogige cactusmussen zijn duidelijk in het nadeel, hun ogen zijn al van verre zichtbaar en ze worden vaak al op jeugdige leeftijd door roofvogels gegrepen. Toch verdwijnen ze niet uit de populatie want dan zou het o-gen moeten verdwijnen. En dát gebeurt niet omdat groenogigen, die ook een o-gen dragen, zeer succesvol zijn.[107] Elk van de drie mogelijke genencombinaties, OO, Oo en oo, noemt men een genotype. We hebben in dit geval dus te maken met drie genotypen.
We zouden ons populatiegenetische onderzoek aan de cactusmus kunnen uitbreiden door behalve naar oogkleur ook naar ooggrootte te kijken. Stel dat er wat dat betreft twee vormen in de populatie voorkomen: grootogige en kleinogige, en dat ook dat verschil door twee genen op één locus wordt bepaald: GG- en Gg-individuen zijn grootogig en gg-individuen kleinogig. Ons onderzoek strekt zich dan opeens uit over negen genotypen: OOGG; OOGg; OOgg; OoGG; OoGg; …; enzovoort. Aard en aantal van de genotypen die we in een populatie onderscheiden hangen dus af van de kenmerken die we bestuderen.
Het woord genotype wordt ongelukkigerwijze in verschillende betekenissen gebruikt. Soms bedoelt men met ‘het genotype van een individu’ de combinatie van genen op een of een paar loci van het betreffende individu. Welke loci men daarbij op het oog heeft blijkt meestal wel uit de context. Maar in zeer algemene beschouwingen is die context niet altijd meer zo duidelijk. Wie beweert dat het fenotype van een individu het product is van zijn genotype en zijn omgeving, heeft daarbij niet een bepaald gen of een paar genen op het oog, maar het totale genetische materiaal van dat individu. Misschien zou het daarom beter zijn om hier het woord genoom te gebruiken: het hele genenbestand van een individu.[108]
Het lijkt me niet zo zinvol om je van een genotype (of een genoom), in de betekenis die ik zojuist suggereerde, af te vragen of het al dan niet altruïstisch is. Waarschijnlijk bedoelen biologen die het over een altruïstisch genotype hebben dat ook niet. Populatiegenetici gebruiken het woord genotype meestal in een andere betekenis. En die is hier wel bruikbaar. Wanneer zij spreken over het verdwijnen van een genotype bedoelen ze niet dat een individu op een bepaald locus opeens geen genen meer heeft. Ze bedoelen dat individuen met een bepaald genotype niet meer voorkomen. ‘Genotype’ verwijst dan dus naar een verzameling individuen die in relevante opzichten – en wat relevant is bepaalt de onderzoekscontext – genetisch gelijk zijn. Ik zal het woord ‘genotype’ in het vervolg zo veel mogelijk in die betekenis gebruiken. Een altruïstisch genotype is dan een genotype waarvan de leden zichzelf (gemiddeld) ten voordele van de leden van een ander genotype benadelen.
Op eenzelfde manier zou je over een altruïstisch gen kunnen spreken: een gen waarvan de dragers zichzelf op de een of andere manier (gemiddeld) tekort doen. ‘Gen’ is dan het ‘kleinst’ denkbare genotype. Maar het is de vraag of bijvoorbeeld iemand als Dawkins dat op het oog heeft wanneer hij het over altruïstische en egoïstische genen heeft. Het is mogelijk dat hij daarbij niet aan een verzameling individuele organismen denkt maar aan een specifiek stukje DNA. Ik zal geen bij die interpretatie passend bioaltruïsmebegrip uitwerken. Je kunt alleen maar in zeer overdrachtelijke zin spreken over het ‘gedrag’ van een stukje DNA, en dat past niet in de rest van mijn betoog. En wie die overdrachtelijkheid accepteert zal er straks weinig moeite mee hebben om alsnog een voor losse genen bruikbaar altruïsmebegrip uit te werken.
Voor fenotype geldt ongeveer hetzelfde als voor ‘genotype’. Soms gebruikt men het voor het totaal aan kenmerken van een individu. Naar analogie van ‘genoom’ zou je hier het woord fenoom kunnen gebruiken. Meestal echter verwijst ‘fenotype’ naar één (‘groene ogen’)[109] of een paar (‘grote groene ogen’) kenmerken van een individu, en in populatiegenetische verhandelingen verwijst het vaak naar alle individuen met een of meer bepaald(e) kenmerk(en). In het vervolg zal ik het in die laatste betekenis gebruiken.
Een voorlopige lijst van mogelijke ‘gedragers’ in een altruïsmedefinitie ziet er dus als volgt uit: individuele organismen, genotypen en fenotypen (de beide laatste opgevat als verzamelingen van individuele organismen).
3.2.2.1.2. gedragingen en strategieën
De volgende variabele in ons raamwerk voor altruïsmedefinities is het gedrag. Dat kan namelijk op verschillende manieren worden gekarakteriseerd.
Wat een dier nu doet is meestal anders dan wat het zo net deed en weer anders dan wat het zo meteen doen zal. Zo bezien is gedrag iets waar je niet over praten kunt zonder dat het intussen al weer veranderd is. Dat is niet alleen voor biologen lastig. En het is niet vreemd dat we daar al in het dagelijks taalgebruik een impliciete oplossing voor hebben. Als we praten over iemands gedrag dan bedoelen we vaak niet wat de besprokene op dit of dat moment precies doet, maar iets algemeners. Als het gaat om iemand die we kennen dan hebben we als het ware een overzicht in ons hoofd van wat voor haar kenmerkende bezigheden en kenmerkende reacties op allerlei situaties zijn. Je zegt van iemand die de hele winter geschaatst heeft niet dat haar gedrag veranderd is wanneer ze ’s zomers regelmatig op de fiets zit. Zeker niet als je weet dat zomerse fietstochten altijd al haar lust en leven waren. Pas wanneer ze onder dezelfde omstandigheden, mooie zomerse dagen, iets anders doet dan voorheen heet haar gedrag ‘anders’.
Biologen hebben hetzelfde idee (denk bij gedrag niet aan wat een organisme op een willekeurig moment doet, maar aan wat voor dat organisme kenmerkende gedragingen onder allerlei specifieke omstandigheden zijn) expliciet uitgewerkt in hun ‘strategie’-begrip.[110] Een strategie is een (volledige) verzameling regels die specificeren wat een individu onder alle denkbare omstandigheden doen zal. In die betekenis kan een strategie als onderdeel van een beschrijving van het fenoom van een organisme worden opgevat.[111]
Het strategiebegrip zoals ik dat hier definieerde staat voor het totale gedrag van een individu. Maar biologen hebben het vaak over strategieën in een ruimere betekenis. (Daarmee bedoel ik: een betekenis waarin er méér dingen onder vallen dan in de eerder gegeven betekenis. Zo is ‘kleine kinderen’ een begrip met een ruimere betekenis dan ‘kleine meisjes’. Alle kleine meisjes zijn kleine kinderen, maar niet alle kleine kinderen zijn kleine meisjes.) In de speltheoretische modellen die met name door Maynard Smith[112] in de biologie werden geïntroduceerd spelen verschillende zogenaamde ‘strategieën’ tegen elkaar. Het bekendste en ook wel zo ongeveer eenvoudigste voorbeeld van een speltheoretisch model is dat van de ‘haviken’ en de ‘duiven’. ‘Haviken’ en ‘duiven’ zijn soortgenoten die zich uiterlijk niet van elkaar onderscheiden maar die zich in bepaalde omstandigheden verschillend gedragen. ‘Haviken’ proberen bij elke ontmoeting met een soortgenoot een gevecht aan te gaan, terwijl ‘duiven’ hun soortgenoten ongemoeid laten en als ze worden aangevallen onmiddellijk weglopen.[113] Op welke manier de individuen in kwestie aan hun eten komen, hoe ze voor hun kinderen zorgen, wanneer en hoe ze hun nest bouwen of hun hol graven, als ze dat doen, blijft buiten beschouwing. ‘Havik’, of ‘duif’, staat dus niet voor een complete gedragsbeschrijving, maar voor een verzameling regels die specificeren welke van een bepérkt scala van gedragingen een individu onder welke van een bepérkt scala van omstandigheden vertonen zal. In die ruime betekenis zal ik de term ‘strategie’ verder gebruiken. De strategie in de betekenis die ik eerder schetste, het hele gedragsfenoom, valt daar ook onder als het grensgeval waarbij het complete scala van gedragingen en het complete scala van omstandigheden zijn gespecificeerd.
In evolutionair perspectief zijn strategieën interessanter dan losse gedragingen. Maar biologie is meer dan evolutiebiologie. Ik zal daarom straks zowel voor strategieën als voor losse gedragingen altruïsmebegrippen uitwerken.
3.2.2.1.3. fitness, kindertal(verm) en welstand
Altruïsme wordt meestal gedefinieerd in termen van fitnesseffecten. Maar, zoals ik al zei, daarnaast kijkt men soms ook naar levensduur, vruchtbaarheid, of vager zaken die je met welstand of welbevinden zou kunnen aanduiden. Aangezien we fitness kunnen opvatten als het product van levensduur en vruchtbaarheid zal ik die twee verder niet apart onderscheiden. Op welstand en welbevinden kom ik straks nog terug. Eerst moeten we het fitnessbegrip zorgvuldiger bezien.[114]
De ‘fitness’ van een individu wordt vaak opgevat als zijn feitelijke aantal nakomelingen, maar volgens John Beatty[115] sluit die interpretatie niet aan bij wat biologen zich verder intuïtief bij ‘fitness’ voorstellen. Om dat duidelijk te maken geeft hij een voorbeeld van twee identieke tweelingbroers van wie de ene op jeugdige leeftijd, tijdens een gezamenlijke wandeling, door de bliksem wordt getroffen en overlijdt, terwijl de ander volwassen wordt en kinderen krijgt. Overwegend dat het ongeluk net zo goed de andere broer had kunnen treffen zullen de meeste biologen aarzelen om de jonggestorvene een fitness van nul en de ander een relatief hoge fitness toe te kennen.[116] En er is een simpele manier om aan die intuïtie recht te doen: definieer fitness niet als het feitelijke aantal nakomelingen van een organisme maar als het aantal nakomelingen dat het gezien z’n geaardheid naar verwachting zal krijgen in de omgeving waarin het leeft.[117] Dat sluit aan bij de gebruikelijke gedachte dat de fitness van een individu een kwaliteit is die hem als het ware tot een bepaald aantal nakomelingen voorbestemt.[118] De fitness van een individu kan bij die interpretatie gebruikt worden om zijn feitelijke nakomelingental te verklaren.
Ik zal zo meteen het verschil tussen het feitelijke aantal nakomelingen van een individu en de fitness van dat individu proberen duidelijk te maken. Maar voor ik dat doe moet ik iets meer zeggen over een begrippenpaar dat voor die uitleg van belang is. Ik zal onderscheid maken tussen de omgeving waarin een individu leeft en de omstandigheden waaronder het leeft. Beide termen hebben betrekking op het totaal van wat zich rondom dat individu bevindt. Alleen, bij ‘omgeving’ denk ik aan de aspecten van dat totaal die relatief constant zijn, terwijl ik bij ‘omstandigheden’ denk aan aspecten die van moment tot moment kunnen veranderen.[119]
Stelt u zich een individu voor dat geboren wordt in een bepaalde omgeving, dat daar opgroeit en daar uiteindelijk overlijdt. Dat individu heeft dan van alles en nog wat meegemaakt, en veel of juist heel weinig nageslacht gekregen. Wat er allemaal precies gebeurd is, hangt af van de omstandigheden waaronder dat individu geleefd heeft. Als die omstandigheden toevallig net iets anders waren geweest had zijn leven heel anders kunnen verlopen. Kortom, de feitelijke levensweg van een individu is er één uit vele mogelijke levenswegen. Je zou je het leven kunnen voorstellen als een wandeling langs een boomvormig vertakt padenstelsel. Op elke splitsing bepalen de aard van het individu en de omstandigheden die hij dán toevallig treft welke van de paden die voor hem liggen hij in zal slaan (zie afb. 3.1).

Bij elke mogelijk levensweg hoort een bepaalde kans: de kans dat een individu dat onderaan de boom begint juist die levensweg zal volgen. Als je voor alle mogelijke levenswegen die kansen kent kun je het verwachte nakomelingental van het betreffende individu berekenen. Dat is gelijk aan de som van kans maal kindertal voor alle levenswegen.[120] We kennen dan wat vaak genoemd wordt de ‘darwiniaanse fitness’ van een individu.[121]
Ik had het een stukje terug over steriele werkbijen en hun kinderrijke koninklijke zusters. Tussen werkster en koningin bestaan geen systematische genetische verschillen, en ze leven in dezelfde omgeving. Het verschil tussen beide wordt uitsluitend bepaald door de omstandigheden waaronder ze opgroeiden. Elke werkster had koningin kunnen zijn als ze op een iets ander tijdstip ter wereld was gekomen. Je zou kunnen zeggen dat de levenswegenboom voor elke ‘vrouwelijke’ bij twee hoofdtakken heeft: werkster worden en steriel blijven, of vruchtbaar worden en uitvliegen met de kans te worden bevrucht en veel kinderen te krijgen. Voor de fitness van een individu, zoals ik die hier heb gedefinieerd, is niet van belang op welke van die twee takken het toevallig belandt. Wat telt is het gemiddelde kindertal van alle mogelijke levenswegen. En daaruit volgt dat steriele werkbijen helemaal geen lage fitness hebben. Die fitness is net zo hoog als hij geweest zou zijn als de betreffende werkster wel koningin geworden was. Bijen volgen net als allerlei andere vliesvleugeligen een boeiende alles-of-niets strategie. Dat is alles.
Het is natuurlijk feitelijk onmogelijk om van een bepaald fenoom in een bepaalde omgeving echt een levenswegenboom te tekenen, al was het maar omdat het aantal splitsingen oneindig groot is. En de precieze kans om een bepaalde levensweg af te leggen zal altijd een onbekende grootheid blijven. Maar dat is geen ramp. Fitness is een theoretisch begrip dat z’n belang ontleent aan de rol die het in verklaringen speelt. En al kunnen we de fitness van een individu nooit meten, we kunnen er vaak wel een schatting van maken op grond van bijvoorbeeld het gemiddelde kindertal van een groot aantal individuen met een bepaald fenoom in een bepaalde populatie.[122]
Op basis van de darwiniaanse fitness van het individu kunnen we nu desgewenst de fitness van een genotype of fenotype definiëren als de gemiddelde fitness van de individuen met het betreffende genotype of fenotype.
Ik ga ervan uit dat de kans om een bepaalde levensweg te volgen afhangt van de omgeving en het fenoom van het betrokken individu. Dat betekent dat de darwiniaanse fitness van een individu alleen kan veranderen wanneer de omgeving verandert.[123] Toch spreken evolutiebiologen heel vaak over fitness alsof het een soort kapitaaltje is dat in de loop van het leven van een individu kan toe- en afnemen. Je ziet dat duidelijk aan de spelers in Maynard Smith’s speltheoretische modellen die voortdurend bezig zijn om fitnesspunten te verdienen of te verliezen. En ook, bijvoorbeeld, aan de manier waarop altruïsme vaak wordt gedefinieerd, namelijk in termen van toe- en afnamen van fitnessen als gevolg van bepaalde gedragingen. Dat zou natuurlijk alleen een manier van spreken kunnen zijn, ingegeven door het besef dat ‘precieze’ formuleringen buitengewoon ingewikkeld zullen zijn. Maar er is ook een andere mogelijkheid: wat ze op het oog hebben is niet de ‘fitness’ maar iets dat ik in het vervolg het ‘vermoede kindertal’ of kortweg ‘kindertal(verm)’ zal noemen.
Bij kindertal(verm) denk ik aan de verwachting op een bepaald moment ten aanzien van het uiteindelijke kindertal van een individu, gegeven de wederwaardigheden van dat individu tot dat moment. Als een individu zijn levensweg afwandelt vermindert bij elke splitsing het aantal eindpunten van de levenspadenboom waar hij nog terecht kan komen. Een individu dat al op zeer jonge leeftijd zijn voortplantingsvermogen verliest, doordat hij steriel wordt bijvoorbeeld, kan alleen nog maar op ‘kinderloze’ eindpunten terecht komen. Een individu dat op het juiste moment net toevallig de toto wint en daardoor de middelen krijgt een groot gezin te huisvesten en te voeden kan vanaf dat moment een stuk geruster zijn wat betreft het aantal kinderen dat het na zal laten. Het kindertal(verm) van een individu verandert dus in de loop van het leven (zie afb. 3.1). Het is natuurlijk niet zo makkelijk te bepalen, maar we kunnen wel bij allerlei gebeurtenissen vaststellen of het daardoor toe- of afneemt: wie een territorium verwerft of een prooi bemachtigt heeft op dat moment een hoger kindertal(verm) dan zij gehad zou hebben zonder territorium of voedsel, en op eenzelfde manier zal een ziekte of een fikse verwonding het kindertal(verm) verlagen.[124]
Er zijn een aantal belangrijke verschillen tussen fitness en kindertal(verm). De fitness van een individu ligt al op het moment van de geboorte vast, terwijl het kindertal(verm) voortdurend verandert met de daagse wederwaardigheden van het individu. De fitness van de leden van een populatie is een maat voor de richting die natuurlijke selectie de populatie op zal sturen, het kindertal(verm) is dat niet. Een stijging van het kindertal(verm) van een individu nú impliceert niet dat het kinderrijk zal overlijden, een daling nú sluit een groot gezin later niet uit. Het kan heel interessant zijn om het kindertal(verm) van een individu te volgen. Ik zou mij kunnen voorstellen dat sommige strategieën leiden tot grote schommelingen van het kindertal(verm) terwijl andere dat niet doen. Het is denkbaar dat strategieën met zo’n capricieus kindertal(verm)-verloop kenmerkend zijn voor sociaal levende dieren. Dat op zich zou een boeiende constatering zijn. Maar het is geen constatering ten aanzien van de fitness van de betrokken dieren. Het is dus belangrijk om fitness en kindertal(verm) goed te onderscheiden.
Dieren doen soms dingen die een opmerkelijk aardige indruk maken. Ze stoppen elkaar stukjes voedsel toe, helpen elkaar van een splinter af, of schikken eens een stukje in zodat een ander ook kan zitten. Ons alledaagse altruïsmebegrip is daarbij moeilijk te gebruiken, we weten niet genoeg van de motieven waarmee die schijnbaar aardige dingen worden gedaan. Maar een bioaltruïsmebegrip gedefinieerd in termen van fitness- of kindertal(verm)-effecten is meteen wat erg zwaar geschut. Zelfs als zou blijken dat over zulk gedrag niets bijzonders te melden valt in termen van fitness- of kindertal(verm)-effecten, dan nog zou het gaan om opmerkelijke gestes die best een eigen naam verdienen. Ik stel voor om in die gevallen de term ‘welstand’ te gebruiken. En die term mag gerust wat vaag blijven.
We kunnen bioaltruïsme dus, wat betreft de munteenheid waarin verlies en winst kunnen worden uitgedrukt, definiëren in termen van fitness, kindertal(verm) en welstand. En daarmee is het overzicht van de elementen die in een bioaltruïsmedefinitie voorkomen afgerond.
3.2.2.2. bioaltruïsme: definities
Het gaat bij een definitie van bioaltruïsme altijd om een gedrager, zijn gedrag en de gevolgen van dat gedrag (voor de gedrager en voor andere betrokkenen) gemeten in termen van een of andere munteenheid. En we hielden na de overwegingen in de laatste drie paragrafen voor elk van die elementen een paar mogelijkheden over: drie mogelijke gedragers (individuen, genotypen en fenotypen), twee manieren om gedrag te beschrijven (in termen van gedragingen en van gedragsstrategieën), en drie maten waarmee we de gevolgen van dat gedrag voor de hoofdrolspeler en voor anderen kunnen meten. Daarmee kunnen minstens achttien verschillende definities worden gemaakt (zie afb. 3.2).
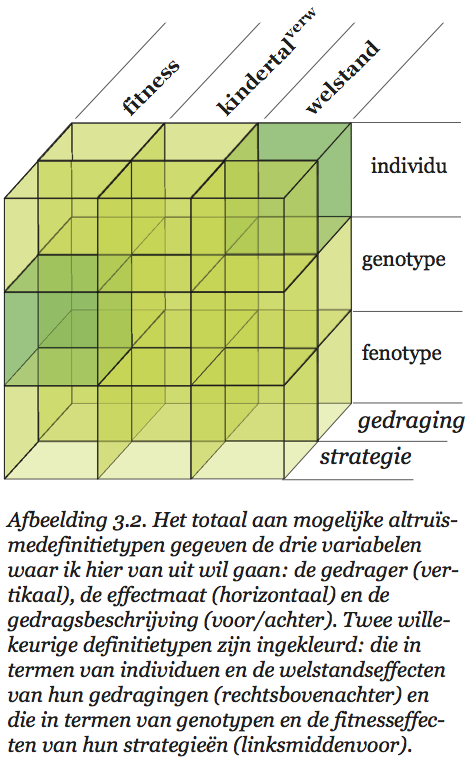
Niet in elke cel van afb. 3.2 zitten zinvolle definities. Sommige combinaties zijn op logische gronden onzinnig, voor andere die niet onzinnig zijn zie ik op dit moment op praktische gronden weinig emplooi. Ik zal nu stapsgewijs laten zien welke cellen ik leeg zal laten (afb. 3.3 vat dat hele eliminatieproces straks nog eens samen), en hoe de definities in de overige cellen eruit zouden kunnen zien.
Laten we beginnen met de achterste helft van de figuur: definities in termen van gedragingen. Het heeft daarbij weinig zin om het over iets anders dan individuen te hebben. Een individu kan zijn buurvrouw een trosje bessen geven, een genotype of fenotype kan dat niet. We kunnen alleen nog kiezen in welke munteenheid we de verlies- en winstrekening opmaken. kindertal(verm) en welstand leveren beide een bruikbare definitie op (definities 1 en 2, zie beneden). Een definitie in termen van gedragingen en fitness geeft meer problemen: losse gedragingen leiden niet tot fitnessveranderingen. Op zich hoeft dat nog geen bezwaar te zijn. We zouden een vergelijking in de definitie kunnen opnemen: het individu in kwestie is niet altruïstisch omdat het zijn fitness verlaagt, maar omdat het (zijn hele leven al) een lagere fitness heeft dan het gehad had als het nu en hier iets anders deed. En dan zijn er in principe twee mogelijkheden. We zouden de levensweg van het individu dat we bestuderen kunnen vergelijken met een andere levensweg uit dezelfde levenspadenboom, of we kunnen het individu dat ons interesseert vergelijken met een (hypothetisch) individu dat een andere strategie volgt. Dat eerste werkt niet. Fitness is alleen gedefinieerd voor de levenswegenboom als geheel. Tussen verschillende wegen binnen een boom kunnen dus geen fitnessverschillen optreden. Het tweede, een vergelijking van verschillende strategieën, brengt ons in de strategieënhelft van het definitieblok. Die komt straks aan de orde (zie definities 3, 4 en 5, beneden).
Definities in termen van gedragingen kunnen dus alleen gaan over individuen en over kindertal(verm) of welstand. We beginnen met de welstand. Een eerste poging: een gedraging A is altruïstisch en het individu I dat A verricht is een altruïst, als ten gevolge van A de welstand van I daalt en de welstand van een ander individu stijgt. Nu zal het heel vaak zo zijn dat er nog meer individuen de gevolgen van A ondervinden. Als die ook van A profiteren is er weinig aan de hand. Maar stel dat sommige anderen ten gevolge van A een verláging van hun welstand ondergaan, dan wordt het toch wel moeilijk om I en A altruïstisch te blijven noemen. Er is gelukkig een eenvoudige manier om dat – conceptuele – probleem op te vangen: neem het individu dat de gevolgen van I’s gedrag ondergaat expliciet op in de definitie, en maak van altruïsme een drieplaatsig begrip.[125] We worden dan vanzelf gedwongen onderscheid te maken tussen het individu tegenover wie I’s gedrag altruïstisch is en alle andere voor wie dat niet geldt:[126]
(definitie 1) Een gedraging A van individu I is altruïstisch tegenover individu J, als I met A de welstand van J vergroot, en zijn eigen welstand verkleint.
Dat is dan in zekere zin zo dicht als je in biologische termen komen kunt bij ons alledaagse altruïsmebegrip. Een definitie in termen van gedragingen en kindertal(verm) is nu ook geen probleem meer:
(definitie 2) Een gedraging A van individu I is altruïstisch tegenover individu J, als ten gevolge van A het kindertal(verm) van J stijgt en het kindertal(verm) van I daalt.
We hebben nu alle mogelijkheden wat betreft losse gedragingen gehad, dus nu zijn definities van bioaltruïsme in termen van strategieën aan de beurt. Ook hier vallen een aantal mogelijkheden meteen af. Een individu dat een bepaalde strategie volgt kan op een bepaald moment zijn kindertal(verm) of zijn welstand verkleinen ten gunste van een ander (en voor fenotypen en genotypen, groepen van individuen, geldt mutatis mutandis hetzelfde). Maar er is een goede kans dat datzelfde individu, met diezelfde strategie, even later zijn kindertal(verm) of welstand ten koste van een ander individu (of wellicht zelfs hetzelfde) zal vergroten. Op grond van de eerste interactie zou je de strategie in kwestie altruïstisch kunnen noemen, maar de tweede interactie geeft je net zo veel reden om die strategie als egoïstisch te karakteriseren. Het probleem is in zekere zin dat de strategie doorgaans niet verandert in de loop van een leven terwijl kindertal(verm) en welstand regelmatig zullen toe- en afnemen. Om die reden acht ik een altruïsmedefinitie in termen van strategieën en welstand of kindertal(verm) niet zo handig. Ik sluit niet uit dat zulke definities kunnen worden opgesteld, maar ik vrees dat ze dan toch zeer gekunsteld zullen zijn. Daarom blijven ze hier verder buiten beschouwing.
We houden nu één groep van combinaties over: definities van altruïsme in termen van strategieën en fitness, waarin de ‘gedrager’ een individu, een genotype of een fenotype kan zijn (zie definities 3 tot en met 5, beneden). Nu zagen we al dat zulke definities niet eenvoudig kunnen zijn. Fitnessen zijn nogal onveranderlijk. Het heeft dus geen zin om je af te vragen of de fitness van, bijvoorbeeld, een individu verandert ten gevolge van zijn strategie. Je kunt alleen het betreffende individu vergelijken met een individu dat een andere strategie volgt. Bovendien hangen de fitnessgevolgen van een strategie af van de omgeving. Die verdient dus ook een plaatsje in de definitie. Dat brengt ons tot een vrij ingewikkeld vijfplaatsig altruïsmebegrip:
(definitie 3) Strategie S van individu I in omgeving E is altruïstisch tegenover individu J vergeleken met de strategie S´ die een (hypothetisch) individu I´ in E zou hebben gevolgd, als de fitness van I in E lager is dan die van I´ in E zou zijn, en als J in E met I een hogere fitness heeft dan hij in E met I´ zou hebben.
Dit altruïsmebegrip is zonder veel moeite ook naar genotypen (definitie 4) en fenotypen (definitie 5) te vertalen:[127]
(definitie 4) Het genotype G waarvan de dragers in omgeving E strategie S volgen is altruïstisch tegenover genotype H vergeleken met genotype G´ waarvan de dragers in E strategie S´ zouden volgen, als de fitness van G in E lager is dan die van G´ in E zou zijn, en als H in E met G een hogere fitness heeft dan het in E met G´ zou hebben.
(definitie 5) Het fenotype F waarvan de dragers in omgeving E strategie S volgen is altruïstisch tegenover fenotype K vergeleken met fenotype F´ waarvan de dragers in E strategie S´ zouden volgen, als de fitness van F in E lager is dan die van F´ in E zou zijn, en als K in E met F een hogere fitness heeft dan het in E met F´ zou hebben.
En daarmee hebben we alle cellen van afb. 3.2 gehad. In afb. 3.3 is het eliminatieproces nog eens kort samengevat.
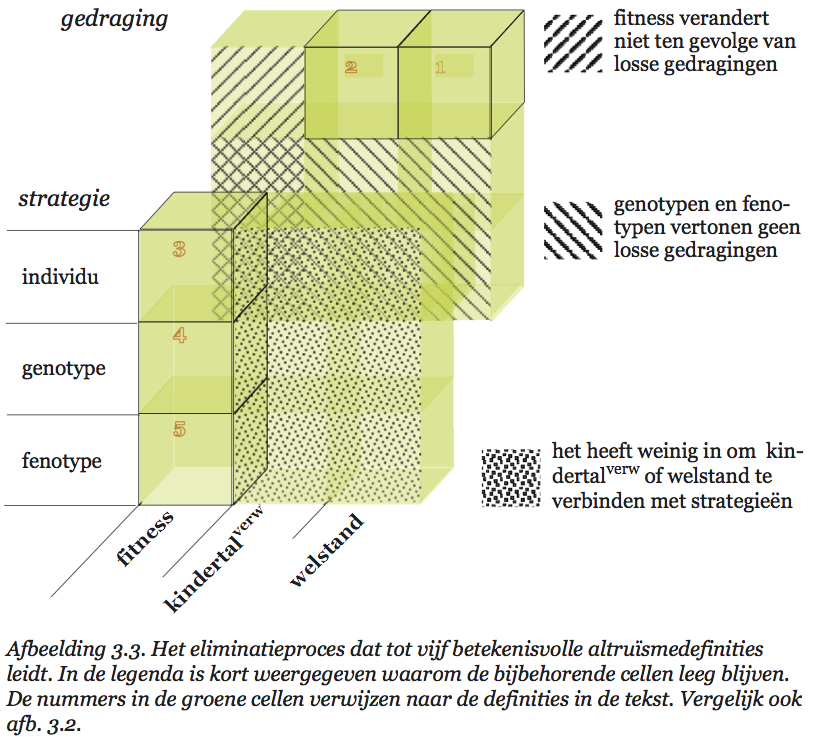
3.2.3. bioaltruïsmedefinities: onderling verband
Altruïsme staat oorspronkelijk tegenover egoïsme. En als het om motieven gaat is dat wellicht begrijpelijk. Je doet iets (primair) voor een ander of je doet iets (primair) voor jezelf. In de sociobiologie zijn beide deel gaan uitmaken van een vierdeling: altruïsme, egoïsme, samenwerking, kwaadaardigheid. En gegeven de typisch biologische definities in termen van verlies en winst voor twee betrokkenen lijkt ook dat een nette indeling op te leveren. Maar zo simpel is het niet.
Eén probleem kwamen we al tegen. Bij bijvoorbeeld altruïsme gedefinieerd in termen van motieven is duidelijk wie het object van de altruïstische daad is, namelijk degene voor wie het positieve effect bedoeld is. Bij bioaltruïsme is dat minder duidelijk. Je zou kunnen zeggen dat zo gauw er iemand profiteert van gedrag waarmee ik mezelf in de vingers snijd er sprake is van bioaltruïstisch gedrag. Maar er zal allicht ook wel een organisme zijn dat nadeel ondervindt van wat ik doe, dus mijn gedrag is al gauw ook ‘biokwaadaardig’. Dat probleem wordt ondervangen door bioaltruïsme niet te definiëren voor één individu maar voor minstens twee. Bioaltruïsme heeft altijd betrekking op een relatie. (En voor de andere categorieën geldt dat evenzeer.) Bij de derde definitie hierboven komt daar nog een derde (hypothetisch) individu bij. En bovendien wordt daar de omgeving cruciaal. Van een nette indeling van gedragingen zijn we dus bij een indeling van vrij ingewikkelde relaties terecht gekomen. Over de relaties tussen die relaties valt meer te zeggen dan ik hier doen wil, maar één punt zal ik hier kort uitwerken.
Een individu dat een ander een deel van zijn eten geeft is duidelijk altruïstisch in de zin van de eerste definitie. Zijn voedselvooraad neemt af en het andere individu krijgt er iets bij. Maar of voedsel delen ook altruïstisch is in de zin van de tweede definitie (in termen van het kindertal(verm)) is minder zeker. We zouden eigenlijk moeten weten wat precies de alternatieven zijn. Als het eerste individu door wat weg te geven een nare ruzie met alle risico’s van kwetsuren en tijdverlies voorkomt, is hij wellicht zo het beste af. Als het ‘geschenk’ volgt op een boze en dreigende blik van de ontvanger ligt het zelfs meer voor de hand die blik egoïstisch te noemen dan de gift altruïstisch.[128] Het is zelfs denkbaar dat de ontvanger zich met zijn gedreig de collectieve woede van zijn groepsgenoten op de hals aan het halen is. En dat zou zijn kindertal(verm) heel wel kunnen verkleinen. Maar als we zulk soort mogelijkheden even negeren en aannemen dat de gever van nu er, wat zijn kindertal(verm) betreft, zonder voedselgift beter aan toe zou zijn en de ander navenant slechter, dan zou er ook in de zin van de tweede definitie van altruïstisch gedrag sprake zijn.
Uit niets volgt echter dat de gever in ons voorbeeld een altruïst is in de zin van de derde definitie. Ik zal met een eenvoudig getallenvoorbeeld laten zien dat altruïsme in de tweede en de derde betekenis weinig met elkaar te maken hebben.[129]
Laten we eens kijken naar dieren met een uiterst simpel leven, een leven waarin maar één keer een cruciale beslissing valt. Bij zulke levens hoort een heel eenvoudige levenswegenboom: een stam met twee takken. Over twee groepen van zulke bomen zal ik het hier hebben. Ze horen bij de individuen die een van twee verschillende strategieën – S1 en S2 – volgen. De bomen verschillen alleen in de kans dat het individu in de linker tak terecht komt. Bij S1 is die kans een half, bij S2 driekwart. Het kindertal in de linkertakken is gelijk, namelijk tien, en dat in de rechtertakken ook, namelijk één (zie afb. 3.4).
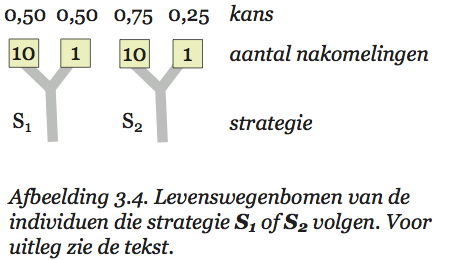
Laten we er eens van uit gaan dat de individuen die S1 of S2 volgen samen met anderen waarvan ik de strategie niet verder invul, in een klein gebied leven waarin geen onbeperkte ruimte voor nageslacht is. Het ligt dan voor de hand dat het gemiddelde kindertal van de groepsgenoten van een individu dat linksaf slaat op de vork in zijn levenswegenboom, lager is dan het geweest zou zijn als dat individu rechtsaf was geslagen. Rechtsafslaan zou je in die situatie altruïstisch kunnen noemen, en wel ‘altruïstisch’ gedefinieerd in termen van het kindertal(verm). Laten we dat kortheidshalve ‘altruïstischverm’ noemen. Individuen die S1 volgen hebben dus een kans van 50% om altruïst(verm) te worden, en voor individuen met S2 is die kans 25%.
De fitness van individuen met S1 is 5,5 en die van individuen met S2 is 7,75. Vergeleken met S2 is S1 dus, in de zin van onze derde definitie, een altruïstische strategie ten opzichte van andere groepsleden.[130] Dat betekent dat individuen met een altruïstische strategie in dit geval een kans van 50% hebben om als niet-altruïst(verm) door het leven te gaan, terwijl individuen met een niet-altruïstische strategie zich nog altijd in 25% van de gevallen als altruïst(verm) zullen ontpoppen. Er is dus geen reden om aan te nemen dat een individu dat zich altruïstischverm gedraagt, ook een altruïstische strategie volgt, en evenmin om ervan uit te gaan dat individuen met een niet-altruïstische strategie géén altruïstischverm gedrag zullen vertonen.
We hebben dus niet te maken met één algemene notie – altruïsme – die zich op verschillende manieren technisch laat uitwerken. ‘Bioaltruïsme’ is een uiterst meerzinnig begrip. En dat is van belang voor discussies over de morele relevantie van sociobiologie. Op dat punt kom ik aan het eind van de volgende paragraaf nog kort terug.
3.2.4. bioaltruïsme en moraal
Sociobiologen wijzen erop dat altruïstisch gedrag veelvuldig voorkomt en ze geven indrukwekkende voorbeelden van zulk gedrag: dieren die hun voedsel delen met soortgenoten, dieren die elkaar waarschuwen bij gevaar, dieren die hun soortgenoten helpen bij nestbouw en zorg voor het nageslacht, dieren die elkaar verdedigen zowel bij ruzies binnen de eigen groep als bij aanvallen van buitenaf.
Sociobiologen menen dat altruïstische gedragingen evolutionair gezien een aparte en extra interessante categorie vormen. Altruïstisch gedrag lijkt op het eerste gezicht een evolutionaire onmogelijkheid. Maar zo simpel ligt het niet. Het is zaak om precies te formuleren wat je op evolutietheoretische gronden wel en niet mag verwachten. Wanneer er in een populatie sprake is van twee of meer strategieën die onderling verschillen in fitness, en als het verschil tussen die strategieën genetisch bepaald is, dan is er reden om te verwachten dat de strategie met de hoogste fitness de andere zal vervangen.[131] Dus wanneer een altruïstische strategie in de zin van de vierde definitie in een populatie blijft bestaan samen met de andere strategie vergeleken waarmee zij altruïstisch is, en wanneer het verschil tussen beide genetisch bepaald is, is er reden om raar op te kijken. Voorbeelden van situaties van dit type ben ik in de sociobiologische literatuur niet tegengekomen.[132]
Kortom, altruïstische gedragingen in de zin van mijn eerste of tweede definitie – in wat je de ethologische zin des woords kunt noemen – komen veelvuldig voor, maar die blijken evolutionair gezien geen enkel probleem te vormen. Ze hebben niets te maken met het eventuele bestaan van genotypen of fenotypen die volgens de theorie weggeselecteerd hadden moeten zijn. En van altruïstische gedragingen of strategieën in de zin van de derde, vierde, of vijfde definitie – de evolutietheoretische zin – die theoretische problemen met zich zouden kunnen brengen, of liever nog: van zulke gedragingen of strategieën sámen met het alternatief ten opzichte waarvan ze altruïstisch zijn, is het bestaan nooit aangetoond. Sociobiologen die altruïsme als een probleem beschouwen – en volgens Wilson is het het meest centrale theoretische probleem voor de sociobiologie – denken kennelijk dat het bestaan van altruïsme in de ethologische zin zou betekenen dat er dingen gebeuren die uit evolutietheoretisch oogpunt een extra probleem zouden vormen. Uit mijn begripsanalyse blijkt dat dat onjuist is.
Dit hoofdstuk begon met de vraag hoe ‘aardig’ je op evolutionaire gronden zou mogen verwachten dat dieren tegen elkaar kunnen zijn. Dat immers was van belang voor de morele relevantie van de sociobiologie. Het antwoord op die vraag lijkt te zijn: heel aardig, maar verwacht niet dat veel dieren zo in elkaar zullen zitten dat ze een ál te lage fitness zullen hebben. Nu denk ik dat er aan al dan niet aardig, zorgzaam, en behulpzaam zijn moreel relevante aspecten zitten (al is moreel gedrag niet hetzelfde als aardig zijn!). Maar morele uitspraken die betrekking hebben op de fitness van individuen kom je maar zelden tegen, zelfs als het om een heel impliciete betrekking gaat. Ook een moraal die in bepaalde situaties altruïstisch gedrag voorschrijft verplicht niemand tot het hebben van een hoge of juist lage fitness. Wat dat betreft hebben evolutionaire theorieën over de mogelijkheid van bioaltruïstische strategieën dus geen morele implicaties. Maar dat betekent nog niet dat evolutionaire theorieën in het algemeen moreel irrelevant zijn. Voor zover morele voorschriften betrekking hebben op de wenselijkheid van bepaalde gedragingen is het heel waardevol om te weten of die gedragingen mogelijk zijn en zo ja onder welke omstandigheden. En iets vergelijkbaars geldt natuurlijk voor moreel onwenselijk gedrag. Op dat punt kom ik in hoofdstuk 5 terug.
Het is verder denkbaar dat je op grond van sociobiologische theorieën uitspraken zou kunnen doen over de wenselijkheid van bepaalde morele voorschriften. De vraag of dat inderdaad kan staat centraal in het nu volgende hoofdstuk.
4 biologie en meta-ethiek
De meeste mensen kennen ethici alleen van de televisie of de radio. Als er moeilijke vragen en beslissingen in het geding zijn – abortus, euthanasie, kernbewapening – verschijnen zij in praatprogramma’s en spreken zich erover uit wat wel en wat niet goed is.
Ethici zijn er in soorten. Sommigen vertellen zonder omhaal wat hoort en wat niet, en daarmee basta. Anderen zijn voorzichtiger. Zij leggen vooral uit waar zoal rekening mee gehouden zou moeten worden, welke afwegingen er in het geding zijn, en in welke volgorde die aan bod zouden kunnen komen, en zijn zo de luisteraar behulpzaam bij diens eigen meningsvorming. Maar allemaal zijn ze op de een of andere manier deskundig als het gaat om vragen van goed en kwaad. Gewone mensen zijn dat kennelijk minder, en dat maakt de vraag interessant waar de kennis van ethici vandaan komt. Volgens sommigen is de sociobiologie een bruikbare bron voor ethische inzichten. In dit hoofdstuk staat die overtuiging centraal.
Gemakshalve zal ik onderscheid maken tussen verschillende takken van de ethiek. Ik houd mij daarbij aan een gebruikelijke indeling.[133] De tak waar de modale tv-kijker het meest mee te maken krijgt is de normatieve ethiek, het vak dat zich zeer direct bezig houdt met de vraag wat wel en wat niet goed is. Ethici die plompverloren komen vertellen wat mag en wat niet, zijn vertegenwoordigers bij uitstek van deze discipline.[134] Het minst zien we de beoefenaars van de descriptieve ethiek, het vak dat waardensystemen en moreel gedrag in verschillende culturen beschrijft. Je zou kunnen zeggen dat het hierbij gaat om sociale wetenschappers die zich gespecialiseerd hebben in normen en waarden. Weer wat vaker, gelukkig, zien we vertegenwoordigers van het derde deelgebied, de meta-ethiek. Meta-ethiek gaat over de grondslagen van de ethiek. Over vragen als: Wat bedoel je eigenlijk als je zegt dat iets (ethisch) goed is? Hoe kom je aan normen? Wat is de relatie tussen verschillende normen en hoe weeg je onverenigbare normen tegen elkaar af? Ethici die niet meteen vertellen wat wel en niet mag, maar die in plaats daarvan zaken aandragen waar je nog niet aan had gedacht en die in je overweging wellicht een rol zouden moeten spelen, maken (een heel praktisch) gebruik van meta-ethische inzichten.
Over normatieve ethiek zal ik het verder nauwelijks hebben. Descriptieve ethiek komt in het volgende hoofdstuk even aan bod. Hier zal het vooral gaan over meta-ethiek, en dan met name over één belangrijke vraag: is het mogelijk om normen, dingen waar altijd maar weer onenigheid over rijst, af te leiden uit (met name sociobiologische) feiten, dingen waarover we het soms makkelijker eens worden, zodat er ten minste enige eensgezindheid kan ontstaan over wat er wel en niet mag.[135]
De gedachte dat evolutiebiologie van belang is voor de meta-ethiek is niet nieuw. Ook vroeger waren er filosofen die in dat belang geloofden (zie 4.2). En wel juist daar waar het gaat om de vraag waar onze meest basale normen vandaan komen. Hun benaderingen worden nu zelden meer echt serieus genomen. Men is het er in ruime kring over eens dat zij smokkelden met het verschil tussen feiten en waarden. Maar de laatste tien jaar zijn er opnieuw pogingen gedaan om uit biologie, en dan met name evolutiebiologie, morele waarden af te leiden, of toch ten minste adviezen over welke waarden we het best als uitgangspunt zouden kunnen nemen. Het tweede deel van dit hoofdstuk biedt van deze pogingen een overzicht.
Eerst zal ik een paar thema’s uit de meta-ethiek kort bespreken,[136] om te laten zien dat biologisch-ethische theorieën ingaan op een serieus probleem.
4.1. meta-ethiek
Wie beweert dat iets goed is gaat er waarschijnlijk van uit dat duidelijk is wat hij of zij daarmee bedoelt. Pas bij verder doorvragen blijkt soms dat verduidelijking niet altijd makkelijk te geven is. Als ik zeg dat ik een goede fiets heb bedoel ik eenvoudigweg dat hij lekker rijdt, dat de rem en de bel het doen en dat hij er niet op de meest ongelegen momenten de brui aangeeft. Mijn fiets is goed omdat hij voldoet aan de voor de hand liggende eisen die ik eraan stel, en die ieder ander aan zijn fiets zou stellen. Dat, en niet meer, bedoel ik dan met ‘goed’. En de bewering dat fietsen goed is is ook vrij makkelijk te interpreteren. Fietsen is gezond, het houdt je in conditie, verdrijft je somberste gedachten, en je komt nog eens ergens. Fietsen is goed omdat het tot gevolgen leidt die mensen wenselijk vinden. Maar wat zou de uitspraak betekenen dat het goed is om een fietsendiefstal te voorkomen, de eerste uitspraak in dit rijtje die we ‘moreel’ zouden kunnen noemen? Is dat voorkómen een handeling die aan bepaalde eisen voldoet? Gaat het er ook hier om wensen van mensen te vervullen? Even lijkt dat zo. De eigenaar van de fiets zal het zeker op prijs stellen dat we de diefstal van zijn rijwiel voorkomen. Alleen, de dief heeft andere wensen, en díe zouden we negeren als we deden wat bijna iedereen goed zou vinden. Sterker nog: het dwarsbomen van de dief is niet zo maar een betreurenswaardige bijkomstigheid, het is in dit geval misschien zelfs wel de essentie van het moreel goede.
Op de vraag wanneer een handeling (moreel) goed is, zijn in de loop der tijden zeer veel verschillende antwoorden gegeven. ‘Een handeling is goed als zij beantwoordt aan de wil van God (of van de goden)’, is een van de oudste en meest eerbiedwaardige interpretaties. Deze interpretatie is natuurlijk nauwelijks bruikbaar voor atheïsten en agnosten. Maar ook gelovige ethici hebben er soms moeite mee. Niet omdat ze twijfelen aan Gods goedheid, maar juist omdat deze interpretatie de uitspraak dat God goed is tot een tautologie maakt. ‘God is goed’ betekent dan zo ongeveer evenveel als ‘God is wat God is’, en dat is geen erg informatieve mededeling. Gods goedheid kan alleen betekenisvol zijn als God niet al per definitie goed is.
Een alternatief zou kunnen zijn: ‘een handeling is goed als zij in overeenstemming is met de morele orde: een stelsel van waarden dat naast of boven de ons omringende fysische werkelijkheid bestaat’.[137] Maar ook die interpretatie is velen niet werelds genoeg.
‘Een handeling is goed als zij leidt tot het grootste gemiddelde geluk voor alle betrokkenen’, is een mogelijkheid die meer mensen aantrekt.[138] Die interpretatie dwingt tot enig rekenwerk, en dat kan op verschillende manieren worden uitgewerkt. Je kunt in elk concreet geval apart de balans opmaken. Zo zou je in ons fietsendiefstalvoorbeeld het verlies van de eigenaar, en eventueel zijn verzekeraar en het overheidsapparaat dat een onderzoek instelt (of zou moeten instellen!), kunnen aftrekken van de winst van de dief, en eventueel die van de heler en de uiteindelijke nieuwe eigenaar, en je ingrijpen afhankelijk stellen van de uitkomst van die berekening. Je kunt ook, in plaats daarvan, proberen vast te stellen of het naleven van de algemene regel ‘voorkom diefstal, als je dat kunt’ in het algemeen het gemiddelde geluk vergroot. Dat is lastiger, maar het lost de morele problemen van de getuigen van een diefstal wel voor eens en altijd op.[139]
Er zijn heel wat ethici die een handeling niet uitsluitend of zelfs maar overwegend op haar effect beoordelen. De bedoelingen van degene die de handeling verricht is volgens hen doorslaggevend. Zij zullen een daad dus alleen goed noemen als die verricht wordt met de bedóeling om een of ander effect te bewerkstelligen.
Weer anderen menen dat ‘deze handeling is goed’ niet meer betekent dan ‘ik zou dit in dit geval doen’, of ‘de meeste mensen zouden dit in dit geval doen’. Morele uitspraken worden dan beschrijvingen van menselijke neigingen. En er zijn ethici die menen dat ‘deze handeling is moreel goed’ helemáál geen feitelijke betekenis heeft. Wie zoiets zegt bedoelt volgens hen, bijvoorbeeld, niet meer dan ‘Doe dit!’, een advies, of een opdracht.
Er bestaan, kortom, nogal wat interpretaties van dat zo vaak gebruikte woordje ‘goed’, en dat geldt voor andere morele begrippen natuurlijk evenzeer. Maar dat hoeft nog niet te betekenen dat we ook van mening verschillen over de vraag welke handelingen goed zijn en welke niet. We zouden het over normatief ethische vragen misschien nog eens kunnen worden. Helaas, echter, blijkt ook dat te optimistisch. In het volgende hoofdstuk kom ik daar op terug.
Sommige meta-ethische interpretaties maken het principiéél onmogelijk om op een voor iedereen bevredigende wijze vast te stellen welke handelingen goed zijn en welke niet. Als ‘Dit is goed’ ‘Doe dit!’ betekent, dan vált er niets vast te stellen.
Bij andere interpretaties is er in elk geval geen sprake van een principiële onmogelijkheid, al zijn er soms wel grote praktische problemen. Misschien bestaat er (zie boven) zoiets als een morele orde die los van, of in elk geval naast, de feitelijke orde der dingen staat. Maar die orde is voor de meeste gewone stervelingen niet erg toegankelijk. En voor Gods wil geldt eigenlijk hetzelfde.
Aan weer andere interpretaties kleeft ook dát bezwaar niet. Je kunt vaststellen of iets je eigen geluk bevordert, of het gemiddelde geluk van de betrokkenen. Er valt achter te komen wat de meeste mensen in een bepaald geval zouden doen. En met wat meer moeite is er misschien zelfs wel achter te komen wat iemand beoogde met zijn gedrag.
Interpretaties van morele termen die het mogelijk maken de morele status van een bepaalde handeling empirisch vast te stellen, noemt men naturalistisch. En naturalistisch-ethische theorieën hebben ontegenzeggelijk het voordeel dat ze duidelijkheid scheppen over de vraag wat wel en wat niet ‘goed’ is. Maar dat voordeel is zeer tijdelijk. Het gaat in de ethiek niet alleen om afstandelijke morele contemplatie maar ook om het afleiden van heel praktische handelingsaanwijzingen. En het zou mooi zijn als die aanwijzingen enig gezag hadden. Maar waar dat gezag vandaan moet komen als je uitgaat van een naturalistische interpretatie van morele termen, is onduidelijk. Wie uit het feit dat een bepaalde handeling volgens een of andere naturalistische interpretatie ‘goed’ is concludeert dat die handeling ook verricht behoort te worden, maakt zich schuldig aan wat vaak ‘de naturalistische drogreden’ wordt genoemd. Een andere manier om dat te verwoorden is: hij negeert het onderscheid tussen ‘zijn’ en ‘behoren’.
4.1.1. de naturalistische drogreden
Stel dat we hebben vastgesteld dat het voorkómen van fietsendiefstallen bijdraagt aan het gemiddelde menselijke geluk, en dat we het daarom goed noemen. Hebben we dan een reden om fietsendiefstallen te voorkomen? Even lijkt dat zo te zijn. Het is immers ‘goed’ om een diefstal te voorkomen. Maar wat zouden we moeten zeggen tegen iemand die fietsendiefstallen niet voorkomt, bijvoorbeeld omdat hij eraan twijfelt of het streven naar menselijk geluk wel zo goed is? Strikt genomen zouden we meteen moeten zeggen dat hij bazelt, immers datgene waarvan hij de goedheid discutabel acht, is per definitie goed! Zijn twijfel is net zo onzinnig als twijfel over de rondheid van een cirkel, of over de vraag of een paar sokken echt wel altijd twee sokken is. Toch zullen velen van ons aarzelen bij de vaststelling dat onze tegenstander een logische fout maakt.
Volgens Moore,[140] de geestelijke vader van onze twijfelaar, kun je na de vaststelling dat het voorkómen van een fietsendiefstal het menselijk geluk bevordert (niet zijn voorbeeld), nog steeds vragen: ‘Maar is het ook goed?’, zonder dat dat een onzinnige vraag is. En hij meent dat je die vraag altijd kunt stellen zolang je werkt met naturalistische interpretaties van morele begrippen. En daaruit volgt dat die de betekenis van morele begrippen kennelijk niet volledig dekken. Wie zulke interpretaties aanhangt maakt zich volgens Moore schuldig aan wat hij de ‘naturalistische drogreden’ noemde.[141] Volgens Moore betekenen morele begrippen altijd méér dan wat zich met een naturalistische definitie vangen laat. Dat ‘meer’ is op zich niet definieerbaar, maar het is wel een heel essentieel deel van de betekenis. Het is juist dát waardoor morele uitspraken redenen geven om iets te doen of te laten. En juist dat ‘meer’ kun je niet zo maar zien en aanwijzen, je kunt het alleen intuïtief ‘zien’, of ‘voelen’, of hoe je dat verder maar noemen wilt. Moore wordt dan ook vaak ingedeeld bij de ethici die men ‘intuïtionisten’ noemt.
Natuurlijk zijn er wel uitwegen voor de ethici die menen dat morele begrippen uitsluitend verwijzen naar feitelijkheden. Ze zouden allereerst kunnen beweren dat wat ze intuïtief zien, als ze zien dat een bepaalde handeling goed is, ís dat die handeling overeenstemt met Gods wil, leidt tot zo veel mogelijk geluk voor zo veel mogelijk mensen, enzovoort. Erg voor de hand ligt die verdediging niet. Ethische naturalisten – en zij niet alleen – verzetten zich vaak juist tegen het intuïtionisme omdat het idee van een intuïtie die ons oordelen verschaft hen niet zo veel zegt.[142] Een andere uitweg zou kunnen zijn dat we een naturalistische definitie opvatten als de definitie van een spiksplinternieuw (feitelijk) begrip ‘goed’ dat we voortaan, bij gebrek aan oude bruikbare begrippen, in morele discussies zullen gebruiken. Maar dat brengt ons weer terug bij een eerder probleem. Wie ervan overtuigd is dat een bepaalde handeling goed (of preciezer: het beste) is zal daar doorgaans uit concluderen dat die handeling ook verricht dient te worden. Dat volgt daar min of meer logisch uit. Maar hoe uit het feit dat een bepaalde handeling tot geluk voor velen leidt, of door God wordt goedgekeurd,[143] direct volgen kan dat zij ook verricht behóórt te worden, valt slecht in te zien. Kortom wie ‘goed’ en ‘kwaad’ definieert in termen van natuurlijke eigenschappen van dingen of handelingen, maakt het die begrippen onmogelijk een doorslaggevende rol te spelen in een morele discussie. De relatie tussen goedheid en wenselijkheid, tussen slechtheid en onwenselijkheid, zou daarmee namelijk verbroken zijn. Je kunt niet tegelijkertijd alle morele begrippen in naturalistische termen definiëren én gefundeerde handelingsaanwijzingen geven. Het is Moore’s verdienste dat te hebben duidelijk gemaakt.[144]
4.1.2. het onderscheid tussen ‘zijn’ en ‘behoren’
Hume beschrijft op treffende wijze zijn ervaring met geleerde werken waarin een ethische theorie wordt gepresenteerd.[145] De auteur van zo’n geschrift begint meestal met verhalen over hoe de wereld in elkaar zit, verhalen waarin het werkwoord ‘zijn’ een belangrijke rol speelt, om dan geleidelijk en haast onmerkbaar over te gaan naar verhalen waarin het werkwoord ‘behoren’ de overhand heeft genomen. Die stap verbaast hem. Er is een belangrijk verschil tussen ‘zijn’ en ‘behoren’, vaak het ‘Is-Ought-onderscheid’ genoemd, waar we niet zo maar omheen kunnen. Men is het er niet over eens of die overgang van ‘zijn’ naar ‘behoren’ volgens Hume logisch onmogelijk is, of niet.[146] Maar één ding is duidelijk: wie die stap wil maken doet er goed aan aan te tonen dat die, in het betreffende geval, ook echt gemaakt kan worden.
In de loop der jaren is het afleiden van voorschriften uit feitelijke beschrijvingen tot een taboe geworden. Wie zich bijvoorbeeld voorzichtig afvraagt of evolutiebiologie ethisch niet tenminste een béétje relevant zou kunnen zijn, krijgt al snel een verontwaardigd ‘Is-Ought fallacy!’ naar het hoofd geslingerd. Dat is jammer omdat zaken waar een taboe op rust alleen al daardoor verleidelijk zijn. Wie iets duidelijk onmogelijks probeert geeft daarmee blijk van onwetendheid of onbegrip, maar wie een intellectueel taboe te lijf gaat toont tenminste moed. Een aantal filosofen hebben naam gemaakt met hun pogingen om uit een feitelijke uitspraak een normatieve uitspraak af te leiden.[147] Maar ze hebben de filosofische gemeenschap niet kunnen overtuigen. Op een van die filosofen – Richards, die uitspraken gebaseerd op evolutionaire overwegingen als uitgangspunt neemt – kom ik straks uitgebreid terug.
4.1.3. feiten en normen
Het minste dat we van Hume en Moore kunnen leren is dat we voorzichtig moeten zijn als we proberen feitelijke en normatieve uitspraken te combineren. En we zullen zo meteen zien dat evolutionaire ethici die voorzichtigheid niet altijd opbrengen. Maar daarmee is niet gezegd dat feitelijke uitspraken geen rol kunnen spelen in morele overwegingen. Wie het zijn plicht acht rekening te houden met de wensen van anderen zal moeten zorgen dat hij weet wat zijn medemensen op prijs stellen. Wie het morele principe huldigt dat niemand verplicht kan zijn tot iets waartoe hij niet in staat is – ‘ought implies can’ (‘behoren’ impliceert ‘kunnen’) is hier de standaardfrase[148] – zal uit het feit dat iemand blind is terecht concluderen dat hem niets verweten kan worden wanneer hij zich voorbij verbodentoegangsbordjes begeeft. En wie de norm aanhangt dat belofte schuld maakt, zal uit het feit dat hij iemand een tientje beloofd heeft terecht afleiden dat hij die ander ook een tientje behoort te geven.[149] Het is heel wel denkbaar dat biologische – en dan met name ethologische – kennis op die manier van belang kan zijn, bijvoorbeeld doordat die ons leert onder welke omstandigheden bepaalde gedragingen de beste, of juist de minste, kans hebben voor te komen. Maar in al die gevallen spelen feiten een rol in redeneringen waarin uit feiten én algemene normen specifiekere normen worden afgeleid. Zonder die algemene normen zouden al die redeneringen drogredenen worden.
4.2. evolutionaire ethiek
Het evolutionaire denken speelt al lang een rol in discussies over de grondslagen van de ethiek. Volgens Comte[150] leidt de evolutie van de mens onontkoombaar naar een uiteindelijk stadium waarin de samenleving als geheel tot volle ontplooiing komt en waarin het individu derhalve vooral door altruïstische gevoelens geregeerd zal worden. En daaruit volgt, zijns inziens, dat altruïsme onze hoogste waarde moet zijn. Ook volgens Spencer[151] leidt evolutie naar een ideaal eindpunt, maar volgens hem is evolutie vooral een kwestie van ‘strijd om het bestaan’ en het ‘overleven van de sterksten’. Zouden we te zeer ons best doen om de minder sterken te helpen in die strijd dan zou dat onze eigen evolutionaire ontwikkeling – en dus het goede zelf – kunnen belemmeren. Altruïsme zal daarom altijd door een gezond egoïsme van de sterksten moeten worden ingeperkt.[152] Voor deze en andere filosofen zijn dus de richting en het eindpunt van de evolutie – en dat zijn feitelijke zaken – normgevend.
Ook nu zijn er weer wetenschappers die de evolutiebiologie willen gebruiken om ethische problemen te lijf te gaan. Van hen is Wilson de meest bekende. Volgens hem is het de hoogste tijd om de ethiek te ‘verbiologiseren’.[153] Alleen de (socio)biologie zal een goede verklaring kunnen geven voor het feit dat wij morele wezens zijn, en voor wat we als moreel gedrag beschouwen. Zonder een gedegen kennis van de biologie zijn onze morele beslissingen slagen in de lucht. En – althans zo lees ik Wilson – veel meer dan die kennis hebben we niet nodig.
Moraal is volgens Wilson uiteindelijk uitsluitend een kwestie van gevoelens. En die gevoelens ontspruiten in de hypothalamus en het limbische systeem.[154] Daar ligt (fysiologisch) vast tot hoeveel altruïsme wij geneigd en in staat zijn. En dat op zich kunnen we weer evolutionair verklaren.[155] We zagen in het vorige hoofdstuk dat het verklaren van onze altruïstische neigingen (in een ethisch relevante zin des woords) lastiger is dan Wilson lijkt te denken. Maar dat is wellicht een overkomelijk probleem. Belangrijker is in dit verband dat Wilson goed en kwaad, althans in zijn vroege werk, kennelijk interpreteert als dat wat mensen wel, respectievelijk niet, willen. Daar is op zich weinig tegen in te brengen, maar het betekent wel dat het morele ‘extra’ waar Moore op wees buiten beschouwing blijft. En dat betekent dat de weg naar een normatieve ethiek zo goed als afgesloten is. Tenzij Wilson kans ziet om uit feiten normen af te leiden. Erg helder en expliciet is hij over dat probleem niet.[156] Maar hij suggereert wel dat we meer kunnen doen dan verklaren alleen.
In zijn boek over de menselijke natuur[157] schetst Wilson het dilemma waarin we belanden als we eenmaal ten volle begrijpen welke richtlijnen voor gedrag in onze hersenen zijn vastgelegd, en hóe die zijn vastgelegd. Die kennis zal ons, volgens hem, in staat stellen om onze eigen toekomst bewust en effectief te sturen. De vraag wordt dan wel welke van die aangeboren neigingen we zullen volgen en welke we zullen intomen. Met een beetje intuïtie komen we wellicht een eind, maar alleen echte, en met zeer veel moeite verkregen empirische kennis van onze biologische natuur zal ons in staat stellen optimale keuzes te doen uit de beschikbare mogelijkheden.[158] Kennelijk valt er te kiezen en ligt de uitkomst van die keuze niet al ‘biologisch’ vast, anders zou het weinig zin hebben om over een ‘keuze’ te spreken, en daar gewichtig over te doen. Maar het vervelende is, en dat negeert Wilson ten enen male, dat je voor een keuze niet alleen kennis maar ook criteria nodig hebt. Waar die vandaan moeten komen blijft in het duister. Onze keuze dient ‘optimaal’ te zijn. Daar zal niemand tegen wezen, zeker zo lang niet is ingevuld met betrekking tot wát die keuze optimaal moet zijn.[159] Wilsons criterium laat zich raden. Hij maakt zich regelmatig zorgen over het voortbestaan van de mensheid. Kennelijk ziet hij dat als een groot goed. En misschien vinden we dat allemaal wel. Maar daaruit volgt nog lang niet dat het ook onze hoogste, of zelfs enige, waarde moet zijn.
Wilson schetst een aantal verklaringen voor het bestaan van homoseksualiteit. Hij suggereert dat homoseksualiteit biologisch gezien waarschijnlijk heel normaal is, al zal het zeldzaam blijven. Daaruit concludeert hij dat een morele veroordeling van homoseksualiteit verkeerd (in de zin van ‘immoreel’) is.[160] Dat is natuurlijk heel aardig van hem, en ik onderschrijf zijn conclusie van harte, maar of het argumént deugt betwijfel ik. Elders[161] stelt hij dat het biologisch heel normaal is dat mannen wat vaker naar seksuele avontuurtjes streven dan vrouwen. De (bij zijn manier van redeneren) voor de hand liggende conclusie dat mannen moreel gezien ook récht hebben op iets meer seksuele vrijheid dan hun vrouwen trekt hij niet expliciet. Daarmee voorkomt hij dat lezers die zich aangesproken voelen door zijn pleidooi voor homo-rechten, nattigheid gaan voelen als er minder emancipatoire idealen in het geding zijn. Kortom, waar Wilson aan normatieve ethiek lijkt te doen, maakt hij zijn verhaal aannemelijk door in te spelen op vage algemene gevoelens,[162] maar logisch gezien klopt er weinig van.
Wilsons eerste pogingen om een biologische basis voor ethiek te ontwikkelen zijn uitgebreid gekritiseerd.[163] En die kritiek was doorgaans nogal vernietigend. Maar uit al die kritiek volgt niet dat sociobiologie irrelevant is voor ethiek. Misschien stelt evolutionair onderzoek ons in staat moraal als verschijnsel beter te begrijpen, wellicht ook kunnen sociobiologische feiten worden gebruikt om uit algemene normen meer specifieke normen af te leiden,[164] en wie weet is het zelfs nog mogelijk om normen af te leiden uit zulke feiten alléén. Al die dingen zijn bepleit door helderder hoofden dan Wilson.[165] En hun analyses verdienen onze serieuze aandacht. Een aantal van deze benaderingen zal ik bespreken. Ik streef daarbij niet naar volledigheid, maar ik zal proberen een representatief overzicht te geven van het brede scala dat nu voorhanden is.
4.2.1. Murphy
Volgens Murphy[166] kunnen we normen rechtvaardigen met meer basale normen, en die weer met nog basalere, maar op een gegeven moment komen we bij onze meest fundamentele normen uit. Die op zich zijn niet te rechtvaardigen. Dat is precies waarom we ze fundamenteel noemen. Maar het feit dat ze fundamenteel zijn, en dus niet gerechtvaardigd kunnen worden, maakt ze in zekere zin ook arbitrair.[167] En dat is voor velen een verontrustende gedachte.
Op het eerste gezicht is er alle reden om ongerust te zijn. Als onze fundamentele waarden inderdaad op niets gebaseerd zijn ontstaat er een kloof tussen de gewichtigheid waarmee we onze morele uitgangspunten belijden, en ons onvermogen om ze te rechtvaardigen. Dat zou het leven tot een vrij bizarre onderneming maken. We spannen ons tot het uiterste in voor waarden die op niets berusten. Het is dan ook niet verwonderlijk dat mensen altijd hebben geprobeerd om hun waarden van een stevig fundament te voorzien. Veel van die pogingen zijn religieus van aard. Maar volgens Murphy werken die niet omdat ze het bestaan van een solide morele basis vooronderstellen, terwijl ze dat bestaan nu juist zouden moeten aantonen.
Murphy onderscheidt twee van zulke benaderingen, maar zijn bezwaar geldt beide. De eerste gaat ervan uit dat de dingen waarnaar we streven in feite Gods doelstellingen zijn. Maar hoe prettig het ook zijn moge om aan Gods zijde te staan, de vraag waaróm Gods doelstellingen goed zijn is hiermee niet beantwoord. Dat lost dus niets op. De tweede benadering gaat ervan uit dat God zelf het goede, of de schepper van het goede, is. Maar om die veronderstelling te toetsen hebben we een onafhankelijke maatstaf voor goedheid nodig, en waar zou die dan weer vandaan moeten komen? Religieuze rechtvaardigingen van morele waarden verzanden dus uiteindelijk in cirkels. En volgens Murphy vergaat het wereldser benaderingen niet beter.[168] Morele waarden hangen uiteindelijk in de lucht, en het leven is in die zin inderdaad absurd.
Murphy meent dat die absurditeit verontrustend is voor iedereen die gelooft dat we voor ons dagelijks leven een met zorg gefundeerd systeem van morele waarden nodig hebben. Als dat geloof terecht is dan zou een ieder die erachter kwam dat zijn ethische principes eigenlijk niet meer dan een luchtspiegeling zijn, in grote nood geraken. Het boeiende is nu dat dat niet gebeurt. Kennelijk is er iets anders dat ons leidt in ons dagelijks leven. Volgens Murphy – en hierin toont hij zich een trouw volgeling van Wilson – worden we uiteindelijk bij elkaar gehouden door onze vermaledijde dierlijke natuur. Onze gemeenschappelijke waarden zijn het product van onze hartstochten en van de gedragspatronen die we vanuit ons evolutionaire verleden hebben meegekregen. Daardoor zullen die ons blijven leiden, ook als we beseffen dat er geen uiteindelijke rechtvaardigingen gegeven kunnen worden.[169]
Dit moreel relativisme betekent overigens niet dat elk ethisch onderzoek verder zinloos is. We kunnen om te beginnen proberen om onze moraal beter te begrijpen. En we kunnen onze ethische systemen verder verfijnen, ze coherenter maken, en er de elementen die zich niet met onze natuur verdragen uit wegwieden. En bij dit alles kan enig inzicht in onze eigen evolutie ons zeer van pas komen. Evolutionaire verklaringen van onze morele uitgangspunten hebben een duidelijk meta-ethisch belang. Al rechtvaardigen ze niets, ze geven in elk geval een goede aanleiding om de juistheid van onze principes nog eens te overdenken. Bovendien is enig inzicht in de menselijke natuur, en dat biedt evolutiebiologie ons, van belang voor normatieve ethiek. Veel ethici, en Murphy met hen, gaan ervan uit dat ‘behoren’ ‘kunnen’ vooronderstelt. Als dat zo is, en als evolutiebiologie ons inzicht verschaft in wat mensen wel en niet kunnen, dan zou dat ons wellicht dwingen bepaalde ethische principes als onverenigbaar met de menselijke natuur te schrappen.
Helemaal overtuigend lijkt Murphy’s verhaal me niet. Hij begint met de stelling dat morele principes een uiteindelijke rechtvaardiging moeten ontberen. Nu is het lastig om overtuigend aan te tonen dat iets onmogelijk is, tenzij het gaan zou om een logische onmogelijkheid, maar daarvan is hier geen sprake. Murphy zou moeten laten zien dat elke poging-tot-nu-toe om een rotsvast fundament voor een ethisch systeem te leggen gefaald heeft. En daarvoor is zijn overzicht te summier. Het zou me ten zeerste verbazen als religieus georiënteerde ethici alleen maar beschikten over de inderdaad nogal bedroevende rechtvaardigingen die Murphy weergeeft.
Laat ik een volstrekt hypothetisch alternatief schetsen waar Murphy’s kritiek geen vat op zou hebben. Stel dat er zoiets bestaat als een door God geschapen morele orde die in principe kenbaar is, maar waar een op zintuigelijke waarneming berustende wetenschappelijke benadering geen toegang toe heeft. Een orde die alleen toegankelijk is voor iemand die bereid is een mystieke weg te gaan. En stel dat een ieder die die weg gegaan is terugkomt met hetzelfde verhaal over hoe die orde eruit ziet. Voor een belijdend gelovige kan dat zeer overtuigend zijn. Maar hoe zou Murphy, die dat duidelijk niet is, reageren?
Het probleem dat hier voor ons ligt betreft de relatie tussen verschillende soorten kennis, en daar doet Murphy, zij het in een iets ander verband, een duidelijke uitspraak over. Hij verwijst naar allerlei orthodoxe dogma’s die in de loop der geschiedenis door wetenschappelijke inzichten werden ondermijnd. De aarde is niet plat meer, en ook niet in zes dagen geschapen, mensen stammen van apen af, en de zon kan nooit voor een beslissende slag om het Heilige Land zijn stilgezet, zulk soort zaken. Nu zijn er volgens hem allerlei manieren waarop je religie tegen dit soort wetenschappelijke ondermijningen kunt verdedigen. Je kunt religieuze uitspraken symbolisch of metaforisch interpreteren, geloof en mysterie tegenover waarneming en rede zetten, of geloof en wetenschap beschouwen als verschillende manieren om over de werkelijkheid te spreken. Maar dat zijn volgens Murphy allemaal ‘ontwijkingsstrategieën’.[170] Hij zal dus wel niet bereid zijn om een mystieke weg te volgen om tot ethische inzichten te komen. De reden daarvoor lijkt te zijn dat dat niet wetenschappelijk is, en dat is het natuurlijk ook niet. Ik zie alleen niet in waarom dat een reden is. Sinds wanneer heeft wetenschap het alleenvertoningsrecht op wijsheid?
Laten we er desondanks eens van uitgaan dat normen inderdaad een uiteindelijke rechtvaardiging moeten ontberen. Volgt daar dan uit dat het leven een zinloze of, zoals Murphy het noemt, absurde onderneming wordt? We moeten ons, denk ik, allereerst realiseren dat normen het droevige lot uiteindelijk in de lucht te hangen delen met feiten. De meest basale feiten kunnen ook niet worden gerechtvaardigd. We zijn het hoogstens over basale feiten wat vaker eens dan over basale ethische uitgangspunten. Maar dat zou een historisch toeval kunnen zijn. Bovendien, wanneer we zeggen dat normen arbitrair zijn moeten we wel beseffen wat we daar precies mee bedoelen. Het is mogelijk dat u en ik uitgaan van verschillende basale normen, en dat geen van ons erin slaagt de ander met redelijke argumenten tot zijn eigen standpunt over te halen. We zouden dan kunnen besluiten elkaars mening te respecteren. Je zou normen, in de situatie die dan ontstaat, ‘arbitrair’ kunnen noemen. Maar dat betekent niet dat mijn normen voor mij om het even zijn, of de uwe voor u. We kunnen allebei in de toekomst van mening veranderen over wat goede morele uitgangspunten zijn, maar we zullen dat doorgaans ervaren als een verdieping van ons inzicht. Alleen, we moeten ons ervan bewust blijven dat wat voor onszelf een goede reden is om van mening te veranderen dat niet ook voor anderen hoeft te zijn. Normen zijn arbitrair in de zin dat u en ik daarover op goede gronden van mening kunnen verschillen, niet in de zin dat we het daar met onszelf niet op goede gronden over eens kunnen zijn.
De absurditeit die Murphy zichtbaar maakt is het gevolg van een spanning tussen het feit dat normen uiteindelijk niet gerechtvaardigd kunnen worden en de vanzelfsprekendheid waarmee we van onze eigen normen uitgaan. De vraag is alleen of die spanning onvermijdelijk is. Zij ontstaat pas als we vol zelfvertrouwen voor ánderen gaan uitmaken wat zij moeten vinden en hoe zij behoren te leven. En daar is geen reden voor. We kunnen best met vol vertrouwen leven vanuit onze eigen normen zonder dat we die ook voor ánderen bindend achten.
Murphy is geen evolutionair ethicus in de eigenlijke zin des woords. Hij gebruikt de evolutietheorie hoofdzakelijk om ons gerust te stellen als de gedachte aan een beetje ethisch relativisme ons benauwd maakt voor een teloorgang van onze beschaving. Ik denk dat we die geruststelling niet nodig hebben. Murphy’s argumenten voor de stelling dat ethische systemen in de lucht hangen zijn niet erg overtuigend, en zelfs als die stelling juist is is er nog geen reden voor bezorgdheid. Wie ontdekt dat zijn morele uitgangspunten geen universele geldigheid hebben hoeft ze nog niet om die reden ook meteen maar helemaal op te geven. Integendeel, die ontdekking kan een goede reden zijn om anderen de vrijheid te gunnen volgens hún normen te leven. Wat dat betreft zou enig ethisch relativisme onze beschaving juist wel eens ten goede kunnen komen.
4.2.2. Ruse en Wilson
De ethische theorie van Ruse en Wilson[171] begint bij twee zaken die volgens hen boven elke redelijke twijfel verheven zijn. Ten eerste: genen hebben het sociale gedrag van dieren goeddeels in de hand. En ten tweede: mensen zijn dieren. Ook óns sociale gedrag staat dus onder genetisch toezicht. En dat heeft, gegeven een beetje evolutiebiologische theorie, een aantal interessante gevolgen. Onder bepaalde voorwaarden, en daaraan is in ons geval voldaan, is het in het evolutionaire belang van genen om hun dragers te laten samenwerken met nauw verwante of minder nauw verwante soortgenoten. Het is zelfs mogelijk dat een individu kinderloos zijn leven geeft voor anderen die dragers zijn van kopieën van zijn eigen genen. We mogen daarom verwachten dat mensen soms, of zelfs vaak, altruïstisch gedrag vertonen.
De auteurs wijzen erop dat altruïsme hier nog niets met goede bedoelingen of met aardig zijn te maken heeft. Of daar sprake van is hangt namelijk af van de manier waarop het altruïstische gedrag tot stand komt. Er zijn globaal genomen twee manieren waarop je een dier altruïstisch zou kunnen maken. Je zou het hele gedragspatroon in de (neuronale) bedrading vast kunnen leggen. Mieren bieden daarvan een aardig voorbeeld. Zij werken, als een ouderwetse automaat, een strak programma af. Op elke gebeurtenis volgt altijd dezelfde standaardreactie, en van bedoelingen is daarbij, naar het zich laat aanzien, geen sprake. Daartegenover staat de wat theoretischer mogelijkheid om een individu te voorzien van zichzelf programmerende superhersenen die alle gedragsmogelijkheden doorrekenen op hun evolutionaire gevolgen en daarna de meest winstgevende mogelijkheid kiezen. Voorbeelden daarvan bestaan eigenlijk niet.
Strakke programma’s zijn riskant. Onder onvoorziene omstandigheden zetten ze je klem. Denk aan de vlieg die net zo lang probeert om door de ruit te vliegen tot ie er definitief bij neervalt. Bij vliegen en mieren is zulk verlies geen ramp, individuen zijn goedkoop en makkelijk te vervangen, maar bij grotere dieren, zoals de mens, zou dat al gauw tot uitsterven leiden. Zo zitten we daarom niet in elkaar. Het andere uiterste, superhersenen, zou ook niet handig zijn. Ze zouden bij elk individu opnieuw gebouwd en geprogrammeerd moeten worden, en dat zou onze toch al lange kindertijd, een periode van grote kwetsbaarheid, nog meer verlengen. Bovendien zouden we waarschijnlijk regelmatig zo lang nadenken dat onze kans om iets handigs te doen al weer voorbij zou zijn.[172]
De oplossing die wijzelf van moeder natuur hebben meegekregen, en wij niet alleen, ligt ergens tussen deze twee extremen in. Wij beschikken over een genetisch bepaald programma van voorwaardelijke gedragsregels en voorkeuren, de epigenetische regels, die onze keuze uit beschikbare gedragsmogelijkheden globaal vastleggen zonder ons op een onwrikbaar spoor te zetten. Die regels vinden we in ons bewustzijn in de vorm van emoties, intenties, terug. Zo, bijvoorbeeld, is onze opréchte hulpvaardigheid de weerslag van de evolutionaire noodzaak om altruïstisch te zijn tegenover verwanten. Moeder natuur heeft ons altruïst gemaakt in de morele zin des woords om te zorgen dat we altruïstisch zijn in de biologische zin des woords in ruwweg die omstandigheden waarin dat evolutionair gezien voordelig is.[173]
Tot zo ver, volgens Ruse en Wilson, de moderne wetenschappelijke verklaring van onze moraal. We zagen inmiddels hoeveel problemen er aan die verklaring zitten.[174] Maar misschien zijn die problemen oplosbaar. Laten we daar alvast een voorschot op nemen en eens kijken hoe onze auteurs een sociobiologische verklaring van moraal gebruiken om een nieuw licht op ethische vragen te werpen.
‘Rechtvaardigt het sociobiologische scenario dat we zojuist schetsten dezelfde morele code die volgens gelovigen door God is gedecreteerd, of die waarvan sommige filosofen geloven dat hij onafhankelijk van de mensheid, net als een wiskundige waarheid, bestaat?’, zo vragen Ruse en Wilson zich af.[175] Die vraag vooronderstelt dat een wetenschappelijke verklaring iets kán rechtvaardigen. Op dat punt komen ze later terug. Eerst stellen ze ons gerust. Volgens de klassieke sociaal-darwinisten zijn organismen gewikkeld in een eeuwige en bloedige strijd om het bestaan waarin ieder alleen op eigen voordeel uit is. Die visie is, mede dankzij de sociobiologie, achterhaald. Natuurlijk impliceert selectie dat er sprake is van een strijd om het bestaan, maar dat is maar één kant van de zaak. Diezelfde natuurlijke selectie leidde tot het ontstaan van altruïsme en wederzijdse hulp bij dieren die in hechte groepen leven. We moeten alleen niet te veel verwachten. Normaal gesproken zullen we ons leven niet geven voor vreemden als daar niet op de een af andere manier weer winst voor de dragers van ons eigen genetische materiaal tegenover staat. Het echte heiligengedrag waartoe sommige kerken ons oproepen is onmogelijk, of alleen mogelijk onder zeer extreme omstandigheden. Onze globale epigenetische regels werken namelijk alleen in ons genetische voordeel onder niet al te bizarre omstandigheden. Meestal dus, maar niet altijd. Een opvallende uitzondering, zoals moeder Theresa, betekent zeker geen bedreiging voor de sociobiologische verklaring van ons gedrag, ze bevestigt slechts het vermoeden dat we geen superhersenen hebben, zodat we, heel soms, minder efficiënt te werk gaan.
Uit de materialistische verklaring van Ruse en Wilson mogen we van hen niet opmaken dat mensen eigenlijk akelige hypocrieten zijn die alleen maar doen alsóf ze om anderen geven. Onder die omstandigheden waaronder we ons altruïstisch zullen gedragen zullen we dat ook meestal oprecht doen. We geloven serieus in onze morele waarden en plichten, en dat zullen we, dankzij de manier waarop we als gevolg van onze evolutie in elkaar zitten, blijven doen. Sterker nog, we zullen het móeten blijven doen. Ethiek is weliswaar een illusie, maar het is een gemeenschappelijke illusie die werkt dankzij het feit dat we erin geloven.[176] De vraag naar de rechtvaardiging van ethiek is met dit alles niet beantwoord. Die vraag kán volgens Ruse en Wilson ook niet beantwoord worden. Hij is in zekere zin gewoon buiten de orde. Ethiek is een functionele menselijke eigenschap en dat is niet iets waarvoor je een rechtvaardiging geven kunt. Verklaren is alles wat erop zit.
Het lijkt even alsof Ruse en Wilson uit het feit dat ze een verklaring van het bestaan van ethiek kunnen geven, direct afleiden dat de inhoud van die ethiek dús niet gerechtvaardigd kan worden. Maar zo simpel ligt het niet. Ze bespreken de parallel tussen een evolutionaire verklaring van ethiek en een evolutionaire verklaring van wiskunde.[177] Het is duidelijk functioneel dat mensen geloven dat twee plus twee vier is. Volgt daar nu uit dat ‘2+2=4’ een gemeenschappelijke illusie is? Integendeel, zeggen Ruse en Wilson, de waarheid van dat sommetje hebben we juist nodig om te kunnen verklaren dat we erin geloven.[178] Maar twee plus twee is vier, voor alles en iedereen. En dat geldt voor morele voorschriften niet. Als we, zoals termieten, in donkere holen zouden leven en onze overledenen zouden opeten, evenals onze uitwerpselen, dan zouden we begrafenissen en een goed functionerend riool als uiterst immoreel beschouwen. Er is dus geen objectieve, los van onszelf bestaande werkelijkheid waarnaar onze morele uitspraken verwijzen.
Dat moraal niet te rechtvaardigen is, volgt dus uit het feit dat we een ándere moraal zouden hebben gehad als we een ánder soort leven waren gaan leiden. Maar het heeft zin om te preciseren waar ’m dat ‘anders’ precies in zit. Het termietenvoorbeeld is een beetje misleidend. Het eten van uitwerpselen is natuurlijk meer een kwestie van smaak, of van fatsoen, dan van moraal. En voor het opeten van overledenen geldt iets dergelijks, al zitten daar duidelijk wel morele aspecten aan. We beschouwen het vaak als onze morele plicht om de overledenen de laatste eer te bewijzen. Alleen, je hoeft moreel niet zo heel verlicht te zijn om te accepteren dat verschillende mensen dat op verschillende manieren doen. Het is duidelijk dat we ons in andere omstandigheden anders zouden gedragen, maar of we ook van andere hoogste waarden zouden uitgaan is maar helemaal de vraag. Een stelsel van waarden is iets anders dan specifiek gedrag in een specifieke situatie. Dat is nou juist een van de aardige dingen die we van Ruse en Wilson kunnen leren!
Kort en goed: Ruse en Wilson bieden een aanloop tot een schets van een mogelijke evolutionaire verklaring van het verschijnsel ‘moreel gedrag’. Ze concluderen uit die verklaring en een paar zeer discutabele veronderstellingen dat er geen objectieve morele orde bestaat. Misschien hebben ze gelijk als ze het bestaan van zo’n orde ontkennen, maar hun argumenten voor dat niet-bestaan zijn verre van overtuigend. Ze zeggen dat hun verklaring geen rechtvaardiging inhoudt. Dat is mooi en bescheiden. Maar ze adviseren ons wel om serieus in moraal te blijven geloven. En de status van dat advies is onduidelijk. Het kan geen moreel advies zijn, dan immers zou het nergens uit volgen en niet meer zijn dan een slag in de lucht. Maar als praktisch, voorwaardelijk advies (‘mocht u willen dat de zaak blijft draaien dan moet u vooral ...’) is het ook niet zoveel waard. We hebben volgens hen immers geen keus. We zitten zo in elkaar dat we in moraal zullen blijven geloven, met of zonder dringend advies dat te doen. Ruse en Wilson pretenderen te pleiten voor wat zij een ‘echte moraal’ noemen. Maar wat zij presenteren is een moraal die berust op een gedeelde illusie, en die hoogstens ‘echt’ is in die zin dat we er echt in geloven. En daar geloof ik echt niet in.
4.2.3. Campbell
Ook Campbell[179] meent dat normen uiteindelijk niet gerechtvaardigd kunnen worden, maar hij verzet zich tegen het nihilisme waartoe die onmogelijkheid sommige filosofen gedreven heeft. Volgens hem is de situatie vergelijkbaar met die in de kennistheorie. We zullen bepaalde dingen moeten aannemen voor we tot kennis kunnen komen, bijvoorbeeld het – onbewijsbare – bestaan van een fysische wereld buiten ons. En onze kennis zal altijd voorwaardelijk zijn met betrekking tot de veronderstellingen waarop ze berust. Maar als we dat eenmaal hebben geaccepteerd valt er nog heel wat zinnigs te doen. Op eenzelfde manier zullen we onze morele uitgangspunten moeten kiezen voor we een hecht doortimmerd ethisch bouwwerk kunnen opzetten, en de normen in dat bouwwerk zijn uiteraard voorwaardelijk met betrekking tot de zelf gekozen uitgangspunten.
Wat we uiteindelijk nodig hebben, volgens Campbell, is een ethiek waarin op een kritische en zorgvuldige wijze secundaire normen worden afgeleid die ons in staat zullen stellen om onze zelfgekozen basiswaarden te realiseren.[180] Maar daarvoor zullen we allereerst zulke waarden moeten hebben. En hoe komen we daaraan zonder uit te gaan van nog weer basalere waarden om onze keuze te rechtvaardigen? Dat probleem lost Campbell op met behulp van de volgende redenering. De ontwikkeling van een normatieve ethiek heeft weinig sociale en praktische betekenis als de basiswaarden daarvan niet vrij algemeen worden geaccepteerd. En hier kan de evolutietheorie ons helpen. Uit de neo-darwinistische versie van die theorie volgt dat alle organismen, mensen inbegrepen, als ingebouwd doel het overleven van hun genen in volgende generaties hebben. En dat betekent dat het overleven van de mensheid een heel geschikte eerste uitgangswaarde kan zijn.[181] Op zich lijkt dat een aardige redenering. Lógisch is er weinig op aan te merken, al gaat de laatste stap te snel (daar kom ik zo op terug). Maar Campbell gebruikt een tweetal premissen die op zijn minst discutabel zijn. Laten we die eens een voor een bezien.
Een normatieve ethiek heeft, volgens Campbell, pas zin als de basiswaarden daarvan vrij algemeen geaccepteerd worden. Dat lijkt zinnig, maar er zijn heel wat zaken in het leven waar duidelijk morele aspecten aan zitten maar waarvoor het helemaal niet nodig is dat iedereen dezelfde normen hanteert. De manier waarop ik met mijn vrienden omga, wanneer ik ze help of hulp van ze verwacht, met wie ik wel en met wie ik niet op vakantie of naar bed ga, zijn allemaal zaken waar duidelijk morele aspecten aanzitten. En ze zijn van sociaal belang in die zin dat er altijd meer dan één mens bij betrokken is. Maar ze hebben geen directe gevolgen voor onze samenleving als geheel. Er is dus geen enkele reden om te proberen ze in een algemeen geaccepteerd ethisch systeem onder te brengen. Er is in het leven heel wat ruimte voor wat je zou kunnen noemen een persoonlijke ethiek.
Een volgend probleem is dat helemaal niet zo duidelijk is waar de grens ligt tussen het gebied waar die persoonlijke ethiek voldoende is en het gebied waar je een meer algemeen geaccepteerde ‘sociale’ ethiek nodig hebt. Sommige mensen verdedigen voor zichzelf het recht op abortus. Anderen menen dat ze, eenmaal in verwachting, de plicht hebben hun kind ter wereld te brengen en het liefdevol groot te brengen, hoe ongewenst het aanvankelijk ook geweest mag zijn. Een samenleving met aanhangers van beide normen komt in grote problemen als tegenstanders van abortus een voorstander proberen tegen te houden, of als voorstanders van het recht op abortus onder bepaalde omstandigheden zouden proberen een tegenstander tot een abortus te dwingen. Als je zo’n samenleving draaiende wilt houden zul je nauwelijks zonder een hogere-orde norm kunnen. Maar dat zou heel wel een norm kunnen zijn die een ieder respect voor de afwijkende normen van anderen voorschrijft. En dat is géén hogere-orde norm in die zin dat beide lagere-orde abortusstandpunten eruit afgeleid kunnen worden.[182] Gegeven zijn uitgangspunten zou Campbells ethiek dus wel eens een vrij beperkte onderneming kunnen worden die heel wat ethische vragen onbeantwoord laat.
De evolutietheorie leert ons, volgens Campbell, dat elk organisme als ingebouwd doel het overleven van zijn genen in volgende generaties heeft. Nu hebben we de evolutietheorie niet zo nodig om vast te stellen dat veel mensen kinderen willen. Maar er zijn heel wat mensen die dat niet willen en die er ook in slagen om kinderloos te blijven. Het enige dat de evolutietheorie ons leren kan is dat er reden is om te veronderstellen dat veel organismen zo in elkaar zullen zitten dat ze zich, mits geplaatst in een geschikte omgeving, zo zullen gedragen dat ze kinderen krijgen. Maar wie daarbij van een ‘doel’ spreekt gebruikt dat woord wel erg overdrachtelijk. Zo overdrachtelijk dat iemand die van een bewust kinderloos medemens beweert dat ze haar doel gemist heeft, niet méér zegt dan dat zij kinderloos is. Nu, dat wisten we al.
Aan het eind van zijn verhaal doet Campbell een wel erg snelle redeneerstap. Hij leidt uit de uitspraak dat elk organisme streeft naar het overleven van zijn eigen genen af dat wij allen streven naar het overleven van het menselijk genoom.[183] En daarbij maakt hij – impliciet – de stap van ‘ik streef naar het overleven van mijn genen en jij streeft naar het overleven van jouw genen’ naar ‘wij streven allebei naar het overleven van ons beider genen’. Een even dramatische als bekende logische fout.[184] Bovendien is het zeer de vraag of we het overleven van de menselijke soort wel onder alle omstandigheden als onze hoogste waarde zouden accepteren. In hun analyse van Wilsons ideeën over ethiek, waarbij het overleven van de menselijke soort eveneens als hoogste ethische waarde uit de bus komt, geven zowel Ruse als Kitcher[185] voorbeelden van omstandigheden waaronder we er waarschijnlijk voor terug zouden schrikken om de mensheid in stand te houden. Het is denkbaar dat zich een situatie zou voordoen waarin de mensheid alleen toekomst had wanneer een flink deel van de menselijke bevolking onverwijld zou verdwijnen. Niet iedereen zal vinden dat dat massamoord rechtvaardigt.
Kortom, Campbells conclusie dat het overleven van de mensheid ons uiteindelijke doel zou moeten zijn, of ten minste een van die doelen, berust op zeer discutabele premissen en zou bovendien bij enig nadenken wel eens op veel minder enthousiasme kunnen rekenen dan Campbell denkt. En het is ook eigenlijk raar dat hij eerst beweert dat basale normen niet gerechtvaardigd kunnen worden om vervolgens uitgebreid uit te leggen waarom bepaalde normen beter zijn dan andere. Ik kan me niet aan de indruk onttrekken dat hij daar met zichzelf in strijd komt.
4.2.4. Richards
Murphy, Ruse, Wilson en Campbell zijn eigenlijk alle vier van mening dat morele principes uiteindelijk niet gerechtvaardigd kunnen worden. Maar dat betekent niet dat ze overtuigd zijn van de onoverbrugbaarheid van de kloof tussen ‘zijn’ en ‘behoren’. Ruse vermeldt dat die kloof ook binnen de filosofie ter discussie staat, en Murphy vraagt zich in een voetnoot[186] af of het onderscheid tussen het feit dat iets hoort en het geloof dat iets hoort, uiteindelijk wel echt houdbaar is, maar daar laten ze het verder bij. Een van de weinigen die serieus en expliciet proberen om normen af te leiden uit de feiten van de evolutiebiologie is Richards.
Richards[187] is zich ervan bewust dat evolutionaire ethici in het verleden nogal grof te werk gingen. Ze stelden plompverloren dat de wereld zoals zij is goed is omdat zij het product van een evolutionaire ontwikkeling is, of ze gingen ervan uit dat dat waar evolutie uiteindelijk toe leiden zal, om die reden alleen, goed is. Zulk soort simplismen probeert hij zelf met zorg te vermijden. Net als zijn voorgangers bedrijft hij evolutionaire ethiek, maar zijn theorie wijkt op essentiële punten af. Althans dat belooft hij ons.[188] Of hij die belofte waarmaakt zullen we later zien.
Richards’ theorie bestaat uit twee delen: een empirisch deel, dat een verzameling feitelijke uitspraken over de evolutie van de mens en de gevolgen daarvan voor de manier waarop we gedragsmatig in elkaar zitten behelst, en een ethisch deel, waarin methoden zijn uitgewerkt om uit die feitelijke uitspraken normen af te leiden. Dat onderscheid is belangrijk voor een goed begrip van Richards’ onderneming. Hij is er niet op uit om ons een bepaalde moraal op te dringen. Het gaat hem er louter om te laten zien dat het mogelijk is om morele uitspraken af te leiden uit feiten. De feiten waar hij van uitgaat zijn voorlopig. Zouden we ze afwijzen,[189] dan kunnen we het eerste deel van zijn theorie door een ander evolutionair verhaal vervangen en opnieuw het tweede deel gebruiken om tot een moraal te komen. Het gaat dus uiteindelijk om dat tweede deel.
Volgens Richards hebben we onze moraal aan onze evolutie te danken. Ten gevolge van onze evolutionaire ontwikkeling hebben we een aangeboren neiging om mensen die we als groepsgenoten beschouwen altruïstisch tegemoet te treden, en dankzij diezelfde ontwikkeling is het precies die neiging die we ‘moreel goed’ noemen.[190] Dus zowel onze altruïstische neigingen als ons morele oordeel daarover vormen een gemeenschappelijke evolutionaire erfenis. Alleen onder zeer uitzonderlijke omstandigheden kunnen individuen ontstaan die die neigingen missen, maar hun bestaan – Richards noemt ze ‘psychopaten’ – vormt geen bedreiging voor deze evolutionaire generalisaties. Tot zover het empirische deel van de theorie.
Richards beargumenteert vervolgens eerst in algemene termen dat we de grondwaarden van een ethisch systeem wel uit feiten af moeten leiden. Hij onderschrijft van harte dat de stap van feiten naar waarden binnen zo’n systeem uitgesloten is. Maar van waarde naar meer basale waarde terugredenerend kom je op een gegeven moment bij de meest basale waarden uit en waarmee zou je die, en daarmee het hele systeem, kunnen rechtvaardigen? Voor een antwoord op die vraag is het heel belangrijk om te weten wat ‘rechtvaardigen’ precies betekent. Volgens Richards betekent ‘rechtvaardigen’: ‘laten zien dat een uitspraak beantwoordt aan aanvaardbare regels, uitspraken of gebruiken’.[191] Een goed voorbeeld in dit verband is de rechtvaardiging van afleidingsregels in de logica. Uiteindelijk kunnen die alleen maar gerechtvaardigd worden door te laten zien dat ze de redeneringen die een redelijk menselijk wezen zou aanvaarden géldig maken, en de redeneringen die een redelijk menselijk wezen zou verwerpen óngeldig. Het feitelijke redeneergedrag van redelijke mensen rechtvaardigt dus een stelsel van regels. En evenzo zou je morele uitgangspunten kunnen rechtvaardigen door te laten zien dat de praktische morele standpunten van morele menselijke wezens eruit afgeleid kunnen worden.
Als we die mogelijkheid afwijzen, en aannemen dat morele beginselen niet gerechtvaardigd kunnen worden, dan belanden we in een gevaarlijk ethisch relativisme. We zouden geen enkele reden hebben om Franciscus van Assisi een beter mens te noemen dan Adolf Hitler, en niets om een beroep op te doen als we een aanhanger van de laatste willen terugbrengen van de dwalingen zijns weegs. De vraag is alleen of je zo iemand zo kunt overtuigen. Zijn eigen morele oordeel, in elk geval dat over Hitler, zou niet volgen uit de beginselen die we hem proberen aan te praten. En daarmee vervalt zijn reden om ons te geloven.[192] Richards’ enige uitweg is dan nog om onze gesprekspartner als psychopaat te kenschetsen en het verder voor gezien te houden. In dit specifieke geval is dat wellicht zo gek nog niet. Maar als het gaat om iemand met minder obscure morele standpunten – een voorstander van humanitaire hulp aan de contra’s in Nicaragua, of een tegenstander van een economische boycot van Zuid Afrika – dan wordt het toch wel een beetje eng om maar meteen te gaan twijfelen aan zijn morele oordeelsvermogen.[193] Ik denk overigens wel dat Richards gelijk heeft als hij zegt dat een beroep op gedeelde morele oordelen ons laatste houvást is als we iemand voor een bepaald moreel principe willen winnen. Alleen, een rechtváárdiging van dat principe, in die zin dat het stom of slecht zou zijn om niet overtuigd te zijn, is het niet.
Maar Richards heeft meer pijlen op zijn boog. Hij zou het zeker met ons eens zijn dat normen niet uit feiten volgen in de logische zin des woords. Maar er is wel een parallel tussen logische en morele redeneringen. In de logica ‘volgen’ feitelijke uitspraken uit andere feitelijke uitspraken dankzij logische afleidingsregels. Die afleidingsregels maken geen deel uit van de redenering die ze rechtvaardigen. En we zagen net dat ze zelf alleen te rechtvaardigen zijn onder verwijzing naar menselijk gedrag. Bínnen de logica echter staat hun geldigheid niet ter discussie, dat zou ook niet kunnen omdat ze als het ware die logica zelf definiëren. Op een vergelijkbare wijze, onder verwijzing naar de menselijke praktijk, zouden we misschien meta-ethische afleidingsregels kunnen opstellen die een systeem van moreel argumenteren zouden definiëren waarbínnen normen uit feiten zouden kunnen worden afgeleid.
De grote vraag is dus of we een meta-ethische afleidingsregel kunnen opstellen die ons toestaat om normen af te leiden uit louter feiten. Van die regel op zich mogen we niet meer eisen dan dat hij acceptabel is voor de meeste mensen, of voor mensen met een normale morele ontwikkeling, of iets dergelijks. Wie meer wil dan dat zou aan meta-ethische regels eisen stellen die je zelfs aan de grondregels van de logica niet stellen kunt.
Richards gelooft dat het in principe mogelijk is om meta-ethische afleidingsregels op te stellen die de stap van feit naar norm mogelijk maken. Als (hypothetisch) voorbeeld geeft hij de regel ‘beschouw de oproepen van een moreel leidsman als ethisch juist’. Als we die regel zouden accepteren dan zou uit het feit dat de paus een moreel leidsman is en het feit dat hij abortus veroordeelt, de voor ieder geldende norm dat abortus ethisch gezien niet door de beugel kan, afgeleid kunnen worden.
Ik kan me voorstellen dat niet elke lezer deze meta-ethische afleidingsregel aanvaardbaar zal vinden. Hij veronderstelt een volgzaamheid die wat in onbruik is geraakt. Toch is dat het belangrijkste bezwaar niet. Er is iets fundamentelers mis. Je kunt bij een moreel leidsman zowel denken aan iemand die leiding geeft in morele kwesties, als aan iemand die leiding geeft en er (daarnaast) een hoogstaande moraal op nahoudt. Als Richards de eerste betekenis op het oog heeft betekent ‘moreel leidsman’ niet meer dan ‘iemand die zich duidelijk uitspreekt over wat zijns inziens wel en niet mag’. Daar is de paus een aardig voorbeeld van, maar hij is de enige niet. Van dat soort lieden zijn er erg veel, ze zijn het nergens over eens, en we kunnen er allemaal wel een paar aanwijzen wier ‘morele’ oproepen we verafschuwen. Kortom, bij deze interpretatie van ‘moreel’ zal niemand, ook niet de meest volgzame ziel, Richards’ voorbeeld van een meta-ethische afleidingsregel aanvaardbaar vinden. Maar bij de tweede interpretatie van ‘moreel’ ontstaan er ook problemen. ‘Moreel leidsman’ gaat dan zoiets betekenen als ‘iemand met hoogstaande morele opvattingen’. Het moet een genoegen zijn de adviezen van zo iemand op te volgen, maar hoe weet je wie een moreel leidsman is in déze zin des woords zonder dat je al beschikt over een moreel oordeel dat je bij die keuze kan leiden? ‘De paus is een moreel leidsman’ is bij deze interpretatie niet een feit, het is een (moreel) oordeel over ’s pausen morele uitgangspunten. En daarmee houdt deze meta-ethische afleidingsregel op te doen wat hij doen moest: het afleiden van morele oordelen uit louter feiten. En de illusie dat die regel dat zou kunnen berust (even afgezien van ons gebrek aan volgzaamheid) op een cruciale meerzinnigheid. We zullen zo meteen zien dat dat ook geldt voor de meta-ethische afleidingsregel die Richards zelf in zijn ethische theorie wil gebruiken.
Om een stap van feiten naar normen te kunnen maken is het noodzakelijk dat we een beeld hebben van de betekenis van morele termen. Volgens Richards betekent ‘behoren’ zoiets als ‘noodzakelijk gemaakt door redenen ontleend aan een gestructureerde context’. Wat precíes een gestructureerde context is legt hij niet uit, maar hij geeft wel een paar voorbeelden. Na een bliksemflits hoort het te donderen, gegeven een aantal natuurwetten. En als we ten gevolge van onze evolutionaire ontwikkeling zo in elkaar zitten dat het belang van de gemeenschap onze hoogste prioriteit heeft (en dat is zo volgens het eerste deel van Richards’ theorie), dan behoren we ons altruïstisch te gedragen. Dat wil dus zeggen: dan zullen we ons altruïstisch gedragen, als ons daarbij niet te veel in de weg zit. Waar het uiteindelijk op neer komt is dat we met de bewering dat iets behoort te gebeuren, bedoelen dat het zál gebeuren, tenzij het door onvoorziene omstandigheden wordt voorkomen.[194] En de meta-ethische afleidingsregel die Richards voorstelt luidt zo ongeveer: ‘Beschouw de handeling die een bepaalde gestructureerde context noodzakelijk maakt als dat wat behoort te worden gedaan’.
Een ding is duidelijk, gegeven het empirische deel van zijn theorie en zíjn interpretatie van ‘behoren’, heeft Richards gelijk als hij zegt dat we ons altruïstisch behoren te gedragen. Maar daarbij gaat het wel om het louter beschrijvende ‘behoren’ van ‘als ik lakmoes in azijn giet dan behoort het rood te worden’! Als dat niet gebeurt dan zijn we verbaasd. En die verbazing wordt gerechtvaardigd (!) door de ‘gestructureerde context’ van de moderne chemie. Evenzo zouden we gegeven de juistheid van Richards’ uitgangspunten verbaasd zijn als mensen niet altruïstisch waren. Een verbazing die door de gestructureerde context van Richards’ uitgangspunten wordt gerechtvaardigd. Maar hoe daaruit ooit zou kunnen volgen dat we er goed aan doen altruïstisch te zijn, dat we daar wel toe verplicht zijn, dat het immoreel zou zijn dat niet te zijn, kortom hoe een beschrijvend ‘behoren’ ooit een moreel ‘behoren’ zou kunnen worden, blijft een raadsel.
4.3. besluit
Richards is de enige van de hier beschreven auteurs die zonder meer probeerde om eerste normen af te leiden uit de evolutietheorie. Het is hem niet gelukt. Ruse en Wilson, en Campbell, kwamen slechts met dringende adviezen voor een bepaald soort ethiek, althans dat is de meest welwillende interpretatie van hun woorden. Maar ook zij gaven geen redenen die mij echt konden overtuigen. Een evolutionaire ethiek lijkt dus vooralsnog onmogelijk. Hebben we die nodig?
Allen, Murphy daarbij inbegrepen, lijken ervan uit te gaan dat we zonder een goed gefundeerde ethiek in een gevaarlijk ethisch relativisme kunnen verzanden. De angst voor ethisch relativisme van veel evolutionaire ethici verbaast me. Het doet me denken aan de angst van veel traditionele christenen die de ontkerkelijking met lede ogen aanzien: wie zijn geloof verliest leeft daarna maar raak. Die angst komt voort uit de overtuiging dat de mens van nature tot alle kwaad geneigd is, en een geloof behoeft om hem in het gareel te houden. Kennelijk hebben evolutiebiologen die het christendom vaarwel zeiden hun angst voor de juistheid van dat pessimistische mensbeeld behouden. Murphy beperkt zich er goeddeels toe die angst te ontzenuwen: mensen zijn zo slecht nog niet. En voor wie echt bang is dat mensen vooral tot kwaadheid geneigd zijn kán dat een geruststelling zijn. Al zullen we wel moeten bedenken dat de altruïstische neigingen waarmee we volgens sociobiologen gezegend zijn misschien wel heel weinig te maken hebben met het ‘altruïsme’ waar ethici over spreken (zie hoofdstuk 3). Hoe dan ook, Murphy’s bijdrage is nog redelijk zinvol en bescheiden. De anderen gaan allemaal een stap verder. Een stap té ver, zoals we zagen. Bij een speurtocht naar eerste waarden voor een ethisch systeem is de sociobiologie ons niet van erg veel nut.
Maar misschien kunnen we van de evolutiebiologie wel iets leren over onze aangeboren neigingen, over onze ‘natuur’. Zulke kennis is van belang als we ervan uitgaan dat onze morele eisen moeten passen bij onze natuur, als ‘behoren’ inderdaad ‘kunnen’ impliceert. En ook als we daar niet van uit willen gaan kan het handig zijn om te weten op welke manier morele doelstellingen het makkelijkst gerealiseerd kunnen worden. Als je diefstal afwijst maar weet dat mensen er nauwelijks weerstand aan kunnen bieden iets mee te nemen dat ze zeer begeren en dat voor het grijpen ligt, dan komen adviezen om die verleiding zo klein mogelijk te maken zeer van pas. De vraag is alleen of evolutiebiologie ons inderdaad zoveel over menselijke neigingen kan leren. Die vraag komt aan de orde in het volgende hoofdstuk.
5 de menselijke natuur
De mens is van oorsprong een steppenjager die leefde in niet te grote groepen van onderling meestal vrij nauw verwante individuen. De volwassen mannetjes in zo’n groep gingen op jacht, werkten daarbij samen met hun groepsgenoten en, zij het in mindere mate, met andere stamverwante groepen. De vrouwtjes deden het huishouden, zorgden voor het kroost en verzamelden wat plantaardig voedsel. Dat patroon ontstond toen onze verre voorouders uit het oerwoud trokken en mens begonnen te worden. Het was ook de enige manier waarop ze zich buiten het oerwoud staande konden houden, en het heeft zich tot op de dag van vandaag, ondanks gewijzigde omstandigheden, in beginsel gehandhaafd. Het is de manier van leven die onze geest heeft gevormd, die gestold is in onze menselijke natuur. Al is de jachtgroep een firma geworden, en de leefgroep een stel vrienden en kennissen – niet zelden de collega’s van meneer. Híj gaat nog steeds op jacht, uitdagingen trotserend om zijn gezin in leven te houden, en zíj blijft nog steeds thuis, soms zelfs als daar geen echte reden voor is. In het belang van kinderen en groepsgenoten zijn ze beiden tot heel wat zelfopoffering bereid, en ook in een iets ruimere kring willen ze nog wel wat hulp verspreiden, dus pure egoïsten zijn het niet. Maar een ethiek waarin alle mensen gelijk zijn kunnen ze hoogstens mooi vinden, ernaar leven is te veel gevraagd. Dat is onmogelijk gegeven de aard van het beestje. Onze aard, de menselijke natuur, is het product van ons evolutionaire verleden. Die aard is bepalend voor het soort ethiek waarnaar we kunnen leven. Daarom is inzicht in onze evolutionaire ontwikkeling onontbeerlijk voor de ontwikkeling van een echt menselijke ethiek.
Dit verhaal kom je in een of andere vorm in heel wat sociobiologische publicaties tegen. Het is de rode draad in Wilsons boek over de menselijke natuur,[195] en we zagen in het vorige hoofdstuk dat ook ethici die evolutiebiologie serieus nemen dat op z’n minst doen omdat ze menen dat evolutiebiologie ons iets kan leren over onze natuur dat relevant is voor een keuze van ethische uitgangspunten. In dit hoofdstuk staat die mening centraal.
Ik zal het in dit hoofdstuk hebben over ‘mensbeelden’ en over de ‘menselijke natuur’. Dat zijn geen synoniemen. Met iemands ‘mensbeeld’ bedoel ik het geheel van iemands ideeën over hoe mensen zijn. Bij ‘menselijke natuur’ denk ik aan een verzameling kenmerken die alle mensen gemeen hebben, of minstens aan de kenmerken die iets in elk geval hebben moet om mens te kunnen zijn. Ik denk dat mensen onderling zeer verschillen, zo zeer zelfs dat ik betwijfel of er over de menselijke natuur veel interessants te melden valt. Dat betekent niet dat ik geen mensbeeld heb. Alleen, in dat mensbeeld is ‘diversiteit’ een belangrijk element.
Ik denk dat evolutiebiologie ons op dit moment minder vertelt over de menselijke natuur dan velen hopen. Er bestaat geen eensgezindheid over ons evolutionaire verleden. Misschien komt die er ooit nog, maar er zijn redenen om daaraan te twijfelen (zie 5.1). Bovendien lijkt het op dit moment erg moeilijk om een reconstructie van onze evolutionaire ontwikkeling te geven zonder uit te gaan van een mensbeeld. En als dan vervolgens uit zo’n reconstructie een beeld van de menselijke natuur opdoemt is dat niet veel anders dan het mensbeeld dat er middels vooronderstellingen eerst is ingestopt. Om dat te illustreren zal ik enkele, onderling zeer verschillende, voorbeelden van reconstructies van ons evolutionaire verleden weergeven en laten zien op welke manier mensbeelden bepalend zijn voor de verschillen daartussen. Mijn voorbeelden zijn niet allemaal even recent, en ze zijn enigszins willekeurig gekozen. Ze laten zien dat (voor)oordelen over de menselijke natuur feitelijk een rol spelen bij het opstellen van evolutionaire reconstructies. Maar daaruit volgt natuurlijk niet dat zoiets ook onvermijdelijk is. Toch meen ik dat er goede gronden zijn om te twijfelen aan de mogelijkheid van een ‘objectieve’ reconstructie. Het bewijsmateriaal dat daarvoor nodig is is schaars en moeilijk te interpreteren. En het is heel wel mogelijk dat dat altijd zo zal blijven. Om dat duidelijk te maken zal ik zo meteen eerst iets meer zeggen over de mogelijkheden en beperkingen van evolutiebiologie.
Een speurtocht naar de menselijke natuur vooronderstelt dat er zoiets als de menselijke natuur bestaat. Maar ook daar kun je vraagtekens bij zetten. Het is onvermijdelijk om zo nu en dan te praten en te theoretiseren over mensen in het algemeen. Maar dat heeft alleen zin als, en voorzover, er karaktertrekken zijn die we (bijna) allemaal delen. Het is zeer de vraag of er veel van zulke trekken zijn. Zowel evolutionaire theorie als onze daagse ervaring geven redenen om te twijfelen aan het bestaan van zoiets als dé menselijke natuur. Uiteraard zijn er allerlei dingen die de meeste mensen gemeen hebben, maar we moeten het belang van die gemeenschappelijke trekjes niet overschatten.
5.1. evolutietheorie en evolutionaire reconstructies
Michael Ruse[196] heeft zich zeer expliciet bezig gehouden met de vraag of er grenzen zijn aan onze kennis van de evolutie. We moeten daarbij volgens hem onderscheid maken tussen (1) de vraag of er zoiets als evolutie heeft plaatsgevonden, (2) de vraag hoe die evolutie dan precies verlopen is, en (3) de vraag welke mechanismen verantwoordelijk zijn voor dat verloop. De eerste vraag gaat duidelijk aan de andere vooraf.
Ik heb dit boek geschreven vanuit de gedachte dat het antwoord op de eerste vraag bevestigend is. Natuurlijk valt dat niet met zekerheid te weten, maar, met Ruse, denk ik dat er nauwelijks een betere verklaring te geven is voor het bestaan van alles wat we om ons heen aan levende wezens en sporen van levende wezens zien.[197] Die vraag komt hier dus verder niet aan bod. De andere zijn relevanter.
Evolutiebiologie kun je opdelen in twee stukken die ruwweg corresponderen met Ruse’s tweede en derde vraag. Aan de ene kant hebben we de eigenlijke evolutietheorie die in algemene termen beschrijft onder welke omstandigheden je wat voor veranderingen kunt verwachten, en aan de andere kant reconstructies van de evolutionaire ontwikkelingen die bepaalde planten- en diergroepen feitelijk hebben doorgemaakt.
Op dit moment gaat evolutietheorie voornamelijk over natuurlijke selectie en genetische veranderingen. We zagen in hoofdstuk 2 dat die theorie op dit moment dan ook nauwelijks in staat is om fenotypische veranderingen ten gevolge van omgevingsveranderingen te verklaren. Maar afgezien van een mogelijke overschatting van de rol van genetische factoren staat niets een verdere ontwikkeling van de evolutietheorie principieel in de weg. Als we aannemen dat factoren die in het verleden tot fenotypische veranderingen hebben geleid dat ook nu nog (kunnen) doen, dan is er alle ruimte om te experimenteren en onze theorie verder uit te werken.
De andere poot van de evolutiebiologie, het reconstrueren van feitelijke ontwikkelingen, worstelt wel met een principieel probleem. Alles wat gebeurt laat aanvankelijk sporen na, maar die sporen worden geleidelijk uitgewist, en komen nooit meer terug. Een voorbeeld: we gaan ervan uit dat giraffen lange nekken en poten hebben omdat ze daardoor voedsel uit hoge bomen kunnen halen. Daarbij veronderstellen we dat de verre voorouders van onze huidige giraffen ooit veel kleinere beesten waren, en dat er een periode was waarin zij te lijden hadden van een zekere schaarste aan voedsel op een hoogte waar ze allemaal makkelijk bij konden. We veronderstellen dat sommige oergirafjes net iets langer waren dan hun soortgenoten waardoor ze toegang hadden tot bladeren die de anderen net niet te pakken konden krijgen. Die net iets langere girafjes hadden dus meer te eten, en daardoor konden ze zich succesvoller voortplanten dan de anderen. Als we nu ook nog aannemen dat lengteverschillen erfelijk waren en dat er regelmatig opnieuw nog weer langere individuen ontstonden die ook weer genetisch van hun soortgenoten verschilden, en dat voedselschaarste een probleem bleef dat opgelost kon worden door iets hoger te grijpen, dan hebben we reden om aan te nemen dat de giraffenpopulatie van generatie op generatie uit steeds en steeds maar weer langere individuen ging bestaan. De evolutietheorie leert ons alleen dat zo’n ontwikkeling zal optreden áls aan de veronderstellingen die ik verwoordde (en misschien nog een paar) is voldaan, maar of daar inderdaad aan was voldaan in het geval van de voorouders van onze giraffen kunnen we van die theorie niet leren. De enige manier om daarachter te komen is een speurtocht naar de sporen van oergiraffen. Misschien vinden we dan wel fossielen van skeletresten die in allerlei opzichten lijken op de botten van moderne giraffen behalve dan dat vele ervan veel korter zijn. Dat zou het waarschijnlijk maken dat we resten van oergiraffen in handen hebben. Wat zouden het anders moeten zijn? En misschien vinden we die resten wel in lagen die verder de sporen dragen van een verslechterend klimaat – droogte, hitte – waardoor het aannemelijk wordt dat er gebrek aan voedsel ontstond. Misschien ook vinden we in die lagen wel opvallend weinig sporen van roofdieren, zodat we extra reden hebben om aan te nemen dat voedsel het belangrijkste levensprobleem voor toen levende planteneters was. Maar als de beesten waarvan we de sporen zoeken geleefd hebben in een gebied waarin fossiliseren erg onwaarschijnlijk was, of als de fossielen die wel ontstonden door latere geologische veranderingen inmiddels weer zijn vernietigd, dan kunnen we fluiten naar aanwijzingen. Veel meer dan zo-had-het-kunnen-gebeuren-verhalen[198] zullen we dan nooit kunnen vertellen. Alleen ons gezonde verstand kan ons nog helpen om zinnige van onzinnige zo-had-het-kunnen-gebeuren-verhalen te scheiden.[199]
Van onze eigen evolutie resten ons wat fossielen en, uit de slotfase, sporen van gereedschapsgebruik. Die op zich bieden niet voldoende houvast om een enigszins compleet beeld van het leven van onze voorouders in de verschillende fasen van onze evolutie te schetsen. Toch worden zulke beelden wel geschetst, en dat betekent dat er heel wat wordt ingevuld op basis van minder directe aanwijzingen. Laten we eens een aantal van die schetsen nader bekijken om te zien hoe dat invullen in zijn werk gaat.
5.2. reconstructies van ons evolutionaire verleden
De interessantste fase in onze evolutionaire ontwikkeling, de fase waarin ons doen en laten wezenlijk begon te verschillen van dat van onze naaste verwanten, begon zo’n acht miljoen jaar geleden toen het Afrikaanse oerwoud ten gevolge van ingrijpende klimaatsveranderingen flink in betekenis afnam.[200] Voor alle oerwoudbewoners betekende die inkrimping een fundamenteel ‘keuze’-moment: ze konden zich terugtrekken in het steeds kleiner wordende woud, of ze konden proberen zich aan te passen aan het nieuw ontstane milieu. De voorouders van wat nu gorilla’s en chimpansees zijn deden het eerste. Maar onze voorouders verlieten het woud en kwamen (uiteindelijk) in opener vegetaties terecht.
Wij zijn behoorlijk veranderd, vergeleken bij wat we ooit als woudbewoners waren. Onze benen werden naar verhouding een heel stuk langer, onze voeten zijn van klim- en grijporganen loopvoeten geworden, we zijn rechtop gaan lopen, hebben een grotere hersenpan – met inhoud – gekregen, onze kaken en ons gebit, vooral onze hoektanden, zijn kleiner geworden, we zijn gereedschap gaan gebruiken, en we verloren de vacht die we ongetwijfeld ooit bezaten. Hoe gebeurde dat, en waarom? Vier antwoorden op die vraag vindt u in de nu volgende paragrafen.
5.2.1.1. de mens als roofdier
Volgens een inmiddels klassieke opvatting, waarvan Morris[201] de bekendste voorvechter is, zijn onze voorouders jagers geworden. Van oorsprong aten ze vooral plantaardig voedsel, vruchten, zaden, bladeren, en zo af en toe een vette rups. Maar in droog open land is van zulk voedsel minder voorhanden. Ze waren dus wel gedwongen om andere voedselbronnen aan te boren. Nu vergt het nogal wat om kleine apen, want dat waren het, succesvol aan de jacht te krijgen. Ze moesten zich daarvoor een aantal typisch roofdierlijke vaardigheden eigen maken. Ze moesten leren om hun prooi te volgen en af te matten, en aangezien ze nooit snel genoeg zouden kunnen worden om dat in hun eentje te doen, moesten ze leren om groepsgewijs te jagen. En ze moesten leren om een prooi te grijpen en om te brengen. Veel veranderingen zijn in dat licht te begrijpen. Lange benen en stevige loopvoeten maakten hen een stuk sneller dan ze waren. Hun groeiende handigheid en intelligentie stelden hen in staat om wapens te maken en effectief te gebruiken, hun naaktheid zorgde voor de broodnodige afkoeling tijdens lange renpartijen. En wat ze in snelheid en dodelijke kracht nog te kort kwamen compenseerden ze door een hechte samenwerking. Nu stelt jagen hoge eisen aan de bewegingsvrijheid. En vooral moeders die met jonge kinderen rondzeulen kunnen aan die eisen niet voldoen. Dat probleem werd in de loop van onze evolutie alleen maar groter doordat kinderen geleidelijk steeds langer afhankelijk werden van de zorg van volwassenen. Onze voorouders waren slimmer dan hun bosbewonende familieleden, maar dat had zijn prijs: kinderen moesten veel meer en veel langer opgevoed en onderwezen worden. De enige oplossing was dus uiteindelijk een strakkere arbeidsdeling. De mannetjes gingen op jacht, zorgden voor het eten, en de vrouwtjes bleven thuis om voor de kinderen te zorgen. En dat had zeer ingrijpende gevolgen.
Voor de thuisblijvende vrouwtjes was het zaak om de garantie te hebben dat manlief weer thuis zou komen met eten voor haar en het kroost. Het risico dat hij elders op de savanne een tweede gezin zou beginnen en daar een deel van zijn jachtbuit zou achterlaten moest ze zo klein mogelijk zien te maken. En híj had vergelijkbare belangen. Als hij zijn vrouw alleen thuis liet, liep hij het risico dat zij zich in een ander zou verlieven, en wie garandeerde dan dat de kinderen die hij thuis gekomen bij haar aantrof zijn eigen kinderen waren? Bovendien moest hij samenwerken met andere mannetjes, en die samenwerking zou door seksuele concurrentie aardig onder druk komen te staan. Het was dus in het belang van de groep als geheel dat de sterkste mannetjes hun zwakkere broeders de mogelijkheid lieten om ook een vrouwtje te verwerven. Het was in ieders belang dat een mannetje en een vrouwtje elkaar trouw zouden zijn. Het monogame huwelijk, dat nu nog kenmerkend is voor de grootste en meest succesvolle culturen, is daardoor ontstaan. Dat huwelijk werd bijeengehouden door een flinke portie samen beleefde seksualiteit. En zeer veel volgens Morris typisch menselijke zaken, van het vrouwelijke orgasme en het voor mensapen zeer forse mannelijke lid, tot het stelsel van regels en taboes dat de seksualiteit omgeeft, zijn zijns inziens in dat licht te begrijpen.
Om van de jacht te kunnen leven moest de mens leren doden. Daar had hij, zwak en relatief tandeloos als hij was, wapens voor nodig. Maar mentaal vergt het ook het een en ander. Een planteneter mist de speciale remmingen die een roofdier ervan weerhouden zijn klauwen en slagtanden tegen zijn eigen soortgenoten te gebruiken. Vanwege zijn groeiende wapenarsenaal moest de mens ook zulk soort remmingen zien te ontwikkelen, anders zou elke kleine ruzie binnen de groep in een bloedbad eindigen. Voor een deel is dat gelukt, voor een deel echter is die aanpassing niet af. Daardoor is de mens de enige diersoort die massale slachtingen onder zijn soortgenoten aanricht.
Kortom, zowel onze mooie kantjes, huwelijkstrouw, een zeer uitgebreide kinderzorg, een groot vermogen tot samenwerking, als onze minder mooie, oorlog en burenruzies met fatale afloop, vinden hun oorsprong in de evolutie van oerwoudvegetariër naar steppejager.
5.2.1.2. de mens als wateraap
Elaine Morgan[202] gelooft niet in bosapen die in één klap steppenjagers werden. Ze heeft daar twee belangrijke redenen voor. In de eerste plaats gaat Morris’ verhaal haar wat te veel over wat er met onze voorváders gebeurde toen zij het bos verlieten. Zij gingen jagen en verloren hun vacht, maar wat voor gevolgen die haaruitval voor vrouwen had, vermeldt het verhaal niet. Die hoefden niet achter wild aan te hollen, ze zouden het dus waarschijnlijk alleen maar koud hebben gekregen terwijl ze thuis zaten huis te houden. En als ze op stap wilden moesten ze hun kleintjes dragen, want die konden zich niet, zoals mensaapjongen, aan hun moeders vacht vasthouden. De voordelen voor hem moeten dus wel erg groot zijn geweest om de nadelen voor haar te compenseren. Daar komt nog bij dat mannen in feite minder vacht verloren dan vrouwen, terwijl het omgekeerde gezien Morris’ verhaal voor de hand zou liggen.
Volgens Morgan is een overgang van planteneter naar grootwildjager veel ingrijpender dan Morris en anderen lijken te denken. De savanne is het leefgebied van groot wild en grote roofdieren. Op groot wild leren jagen kost tijd, en de technieken en vaardigheden die je nodig hebt om je tegen grote roofdieren te verdedigen verwerf je ook niet op een achternamiddag. Onze voorouders zouden zijn verhongerd of opgegeten voor ze hun eerste wapentuig hadden kunnen maken en gebruiken. Volgens Morgan ging de mens dan ook niet direct van oerwoud naar savanne, maar via een tussenfase waarin hij waterbewoner, of minstens kustbewoner werd. In het kustwater is van alles te eten waar je niet meteen als ervaren jager met knots of speer achteraan hoeft te hollen. Voor een beest dat regelmatig rupsen en termieten heeft gegeten – en dat is gebruikelijke kost voor moderne mensapen, dus waarom niet voor onze oerwoudbewonende voorouders – is de stap naar garnalen en ander klein grut niet zo groot. En dan is er tijd genoeg om eerst te leren dat je met stenen schelpen kunt kraken, en daarna dat ook grotere beesten met een welgemikte slag kunnen worden verschalkt. En water is een veilig toevluchtsoord voor wie door een roofdier wordt achtervolgd. Pas veel later, toen ze hun wapentuig hadden geperfectioneerd, en toen het klimaat weer wat beter werd, zouden onze voorouders het water hebben verlaten en het land zijn ingetrokken.
Met dit verhaal kun je heel wat menselijke kenmerken verklaren, ook een aantal kenmerken waar Morris wijselijk niet van rept. In het zeewater konden onze voorouders zich met hun korte kromme beentjes prima redden, maar lange sterke benen waren wel handiger. Je kunt er verder mee van de kant en je kunt er veel beter mee zwemmen. Een vacht is in het water niet zo handig. Echt warm houdt die je niet, en in een milieu waar kinderen vrijwel blijven drijven hebben die aan een beetje hoofdhaar houvast genoeg.[203] Gereedschap voor het doden van prooidieren ligt aan de kust voor het oprapen. Onze voorouders zouden de enigen niet zijn geweest die ontdekten dat je met stenen schelpdieren te lijf kunt. De zeeotter deed dat ook.
Wij verschillen in tal van opzichten van andere mensapen, en voor veel van die opzichten geldt dat we daarin wél op waterdieren lijken. Ik noem er een paar: onze naaktheid, ons onderhuids vetweefsel, de richting van onze (rudimentaire) haren, onze duikreflex, dat wil zeggen het feit dat ons hart, als we onder water verdwijnen, iets langzamer gaat kloppen waardoor we extra zuinig met onze zuurstofreserve omgaan, en onze tranen. Wij zijn de enige mensaap die huilen kan.
Directere aanwijzingen voor een waterig verleden zijn er niet. Er zijn geen ‘menselijke’ fossielen uit de tijd – ruwweg het Plioceen – die we volgens Morgan in het water hebben doorgebracht. Er zijn dus ook geen aanwijzingen dat we in die periode ergens ánders hebben geleefd.
Volgens Morgan zijn onze voorouders aan het eind van het Plioceen weer landinwaarts getrokken. Langs de rivieren in eerste instantie, en later geleidelijk meer en meer van het water af. Ze meent dat dat migratiepatroon terug te vinden is in de fossielen van onze eerste onbetwiste savannenbewonende voorouders, de Australopithecinae. Hoe ouder die zijn, hoe dichter ze bij de (oost-) kust van Afrika gevonden werden. En ze suggereert dat nog oudere fossielen gevonden zullen worden ten oosten van wat nu de oudste vindplaatsen zijn. De toekomst zal uitwijzen of ze daar gelijk in heeft.
Morgan had twee bezwaren tegen het verhaal van Morris en de zijnen. Vrouwen en kinderen spelen er een zeer secundaire rol in en de stap naar het jagerschap is wel erg groot. Er zijn wetenschappers die die bezwaren delen, maar die de stap naar een wateraap niet nodig vinden.
5.2.1.3. de mens als verzamelaar
Tanner,[204] die van huis uit antropologe is, begint haar reconstructie van onze evolutie met een uitgebreide bespreking van het leven van moderne chimpansees. Ze doet dat omdat het onmogelijk is een beeld te ontwikkelen van het doen en laten van onze vroege voorouders zonder daarbij andere diersoorten als vergelijkingsobject te gebruiken. Ze schetst hoe evolutiebiologen, soms zonder opgaaf van redenen, bavianen als uitgangspunt nemen. Volgens Tanner is dat een ongelukkige keuze. Van alle primaten zijn we het nauwst met chimpansees verwant en met hen hebben we het meest gemeen, dus kunnen we van chimpansees ook het meeste leren over hoe onze voorouders ooit geleefd zullen hebben.[205] Om een zelfde reden bespreekt ze uitgebreid het leven van moderne bosjesmannen, mensen die leven in omstandigheden die overeenkomsten vertonen met die van onze verre voorouders, en waarvan de economie waarschijnlijk zeer lang ongeveer hetzelfde is gebleven.
Tanner gaat ervan uit dat onze voorouders al in het woud, of aan de rand van het woud, de eerste stappen hebben gezet naar een dieet dat uit meer dan vruchten en bladeren bestond. Maar volgens haar is de mens pas vrij laat in zijn evolutie gaan jagen. Eerst moesten het gereedschapsgebruik, de onderlinge communicatie en de samenwerking op een aanvaardbaar peil worden gebracht.
De eerste en grootste stap was die van scharrelaars die eten wat hen voor ogen komt naar verzamelaars die ook dingen eten die je niet zo direct in de bomen ziet hangen, die veel gerichter zoeken, daar grote tochten voor maken, en die eten kunnen bewaren, meenemen en delen met anderen. De savanne biedt voedsel genoeg voor een verzamelaar, maar het groeit daar niet allemaal aan bomen. Een deel zit ondergronds of anderszins verborgen: wortels, knollen, insectenlarven, eieren. Nu wroeten ook mensapen wel eens in de grond en als dat iets eetbaars oplevert eten ze dat op. Onze voorouders hoefden die neiging dus alleen maar verder te ontwikkelen. Voor bewaren en delen geldt iets dergelijks.
Het dier dat de savanne introk was al redelijk toegerust om daar voedsel te vinden en zich de roofdieren van het lijf te houden. En daarna had het alle tijd om zich menselijker trekjes te verwerven. Het had regelmatig dingen te dragen, voedsel, een graafstok, het eerste mensengereedschap, dus het had zijn handen veel te vol om daarop te blijven lopen. Het zal daarom steeds vaker en langduriger zijn achterpoten gebruikt hebben.
Volgens Tanner leefden de eerst savannenbewonende mensapen in groepen die bestonden uit vrouwtjes met hun kinderen. Mannen leefden wat aan de rand van zo’n groep. En ze stelt zich voor dat die vrouwtjes met zorg hun partners hebben uitgezocht, zij het voorlopig niet voor zoiets als een vaste relatie. Mannen die zich vrouw- en kindvriendelijk opstelden zouden daarbij in het voordeel zijn geweest. Het uiterlijke verschil tussen de seksen, dat bij chimpansees, onze naaste verwanten, al vrij klein is zou daardoor nog verder zijn verkleind.[206]
Er is een belangrijk verschil tussen het verhaal van Tanner en dat van Morgan. De Australopithecus die aan het eind van Morgans verhaal de savanne in trekt is een naakte aapmens die gewend is vlees te eten en die aan de kust geleerd heeft om met stenen grote dieren te doden. Zijn gereedschapsgebruik is nauw met dat doden verbonden. Tanners Australopithecus is een harige mensaap die vooral plantaardig materiaal eet. Dat betekent dat Tanner zich niet uitlaat over de oorzaken van onze naaktheid. Wat dat planteneten betreft heeft Tanner een aardige troef in handen. Alle Australopithecus-soorten hadden steviger kiezen dan wij nu hebben en die zijn altijd duidelijk flink gesleten. En dat doet vermoeden dat plantaardig materiaal een belangrijk bestanddeel van hun voedsel was.[207] En voor zover zij vlees aten hoeft dat nog niet te betekenen dat ze van de jacht leefden. Het menu van verzamelaars bestaat weliswaar voor een groot deel uit planten, maar bevat daarnaast ook eieren, insecten, kleine zoogdieren, hagedissen en allerlei andere dieren die in principe met de hand gevangen en gedood kunnen worden, zonder dat daarvoor echt hoeft te worden gejaagd.
5.2.1.4. de mens als voedseldeler
Leaky en Lewin[208] komen met een soort synthese van de verhalen van Morris en Tanner. Net als Tanner menen ze dat voedsel verzamelen een belangrijke stap in onze evolutie is geweest, maar ze vinden dat zij het belang van de jacht wel erg onderschat.[209] Volgens hen is het cruciale punt niet zozeer wat onze verre voorouders aten, als wel hoe ze met hun eten omgingen. Zij gingen er als eersten toe over op grote schaal voedsel te delen. Leaky en Lewin gaan ervan uit dat plantaardig voedsel, en dierlijk voedsel dat zich zonder veel geweld laat verschalken, werd verzameld door de vrouwen en dat dat werd gedeeld met leden van het eigen gezin, terwijl de mannen op groot wild jaagden en hun jachtbuit deelden met alle leden van de groep. Net zoals dat nu nog bij jagers-verzamelaars, of liever bij verzamelaars-jagers, gebeurt.
De jacht is volgens Leaky en Lewin geleidelijk ontstaan uit aaseten. Aanvankelijk zouden dode dieren de belangrijkste bron van dierlijke eiwitten geweest kunnen zijn. Veldonderzoek laat zien dat daar op een modale savanne heel wat van voorhanden is, zij het dan dat je er niet op vertrouwen kunt daar voortdurend van te kunnen leven. Maar je kunt zo als het ware al vast voor een deel van vlees leven, en de smaak te pakken krijgen, terwijl je alle tijd hebt om de voor de jacht benodigde technologie te ontwikkelen.
Voor het leven op de open savanne, en zeker in gebieden waar het voedsel schaars is, achten Leaky en Lewin een sociale structuur als die van de daar levende bavianen het meest geschikt. Een mannetje met een harem van wijfjes. In zo’n structuur zijn de mannetjes gevrijwaard van een voortdurende strijd met groepsgenoten om seksuele voorrechten, en de vrouwtjes hoeven niet met mannetjes te concurreren om voedsel. Leaky en Lewin geloven dan ook dat mensen van nature enigszins polygyn zijn.
5.2.2. mensbeelden en reconstructie
Het is duidelijk dat deze verhalen onderling verschillen. De vraag is nu hoe dat komt. Wellicht zijn niet alle auteurs even deskundig.[210] Maar daarmee zijn, denk ik, niet alle verschillen verklaard. Er is een belangrijker oorzaak. Dat zal ik aan de hand van één aspect van die reconstructies toelichten.
Volgens Morris is de stap naar monogamie een oude en cruciale ontwikkeling. Het was een functionele stap, die een noodzakelijke arbeidsdeling mogelijk maakte. En het was een stap die verklaart waarom mensen ook nu hoofdzakelijk monogaam zijn. Morris weet ook wel dat monogamie niet universeel is, maar hij meent dat het wel kenmerkend is voor alle grote en succesvolle samenlevingen, nu en in het verleden. Polygynie (veelwijverij), om over polyandrie (veelmannerij) maar te zwijgen, komt alleen voor in weinigbelovende culturen die als het ware een evolutionair doodlopende zijweg zijn ingeslagen. Monogamie geeft ons de beste levenskansen en is dus kennelijk het meest in overeenstemming met onze ware aard. Velen zullen dat zo intuïtief een rare redenering vinden, en ik kan me dat goed voorstellen, maar het is de vraag of het een zo veel slechtere redenering is als die van Tanner of die van Leaky en Lewin. Voor wie heilig gelooft in monogamie als uiterst menselijke karaktertrek is Morris’ verhaal wellicht heel plausibel.
Leaky en Lewin nemen aan dat polygynie meer in onze aard ligt. Ook zij komen met het argument dat dat een functionele relatievorm was voor een aap die het oerbos verliet. Savannenbewonende bavianen leven immers ook in polygyne groepen. Dat klinkt al wat zakelijker, maar juist deze visie, die ook door bijvoorbeeld Wilson[211] wordt aangehangen, kreeg heftige aanvallen van feministische zijde te verduren. Hij zou louter voortkomen uit vooroordeel en de behoefte om de seksuele dubbele moraal een biologische ‘rechtvaardiging’ te geven.[212] Bij sommige auteurs, Wilson bijvoorbeeld, krijgt dit verhaal wellicht trekken van een rechtvaardiging, maar bij Leaky en Lewin is daarvan geen sprake. En in hoeverre vooroordelen een rol spelen is moeilijk na te gaan. Al is het maar omdat het soms zo moeilijk is onderscheid te maken tussen voor- en andere oordelen. Feit is dat je Leaky en Lewins conclusie een beetje plausibel moet vinden voor hun redenering je kan overtuigen, maar dat geldt voor meer verhalen in deze context.
Tanner schetst de mens, man én vrouw, als lichtelijk polygaam. Bij haar speelt het feit dat ook chimpansees dat zijn een grote rol. Maar wie niet in die polygamie gelooft zal daarin makkelijk een argument vinden om het gebruik van de chimpansee als model af te wijzen. In zekere zin lopen we in cirkels rond. De argumenten van de verschillende auteurs zijn vooral aanvaardbaar voor wie hun conclusies aanvaardbaar vindt. En dat komt doordat het bewijsmateriaal zo indirect is. Kunnen we daar omheen?
Wat weten we nu bijvoorbeeld van onze eigen verre voorouders? We hebben wat fossielen, al zijn dat er vrij weinig vergeleken bij het aantal individuen dat ooit het levenslicht moet hebben aanschouwd. Nu hebben paleontologen een goed ontwikkeld vermogen om uit het bijna niets een heel beest te reconstrueren. Maar zonder een vage voorstelling van het soort beest waarvan je de fossiele resten in handen hebt kun je die fossielen nauwelijks op een zinnige manier interpreteren. Zelfs voor zoiets bescheidens als het in elkaar zetten van een skelet – even aangenomen dat je los verspreide botten gevonden hebt – zul je bepaalde beentjes als bijvoorbeeld ‘typische middenhandsbeentjes’ moeten herkennen, een andere reden om ze niet heel ergens anders op te nemen is er niet. En dat herkennen kan alleen op basis van een analogie met bestaande beesten waarvan je de precieze bouw kent.
Behalve fossielen hebben we ook nog allerlei resten van prehistorisch gereedschap. Maar ook daar kun je minder mee dan men wel eens suggereert. Morris en Wilson zien in elke scherp gepunte steen een wapen en in elke scherpe steenrand een mes om de jachtbuit uit te benen of een schraper om huiden te prepareren. Tanner ziet in datzelfde materiaal graafwerktuigen, en messen om knollen of graafstokken mee te snijden.
Andere directe aanwijzingen zijn er niet. Er zit dus weinig anders op dan een voorzichtig maar vérgaand gebruik van analogieredeneringen. Chimpansees poeren wel eens in de grond dus dat zullen oermensen ook wel gekund hebben. Bij de bosjesmannen jagen de mannen en verzamelen de vrouwen, dus zo zal dat bij Australopithecus ook wel gegaan zijn. In alle succesvolle menselijke culturen was en is monogamie de regel, dus zal het wel een oude menselijke eigenschap zijn. Bavianen die op de savanne leven leven vaak in groepen van een dominant mannetje met zijn harem, dus dat zal de mens toen hij het woud verliet ook wel zijn gaan doen. Onderhuids vetweefsel en een naakte huid zijn typische waterdierkenmerken, dus zullen onze voorouders ook wel waterdieren geweest zijn. Maar als je al die analogieën even serieus neemt kom je uit bij de tegenstrijdigheden tussen de verschillende reconstructies die ik hierboven weergaf. Er zal dus gekozen moeten worden.
We zullen moeten kiezen welke dieren- en welke mensensamenlevingen we als model voor ons evolutionaire verleden zullen gebruiken. En ik zie niet zo goed hoe je die keuze zou moeten maken. Voor elke mogelijkheid zijn wel argumenten aan te voeren. Chimpansees zijn onze naaste nog levende verwanten. Bavianen zijn onze naaste verwanten die, net als onze voorouders ooit, op de savanne leven, en wat dat betreft zijn nu nog levende verzamelaars-jagers ook heel interessant, alleen er zijn een heleboel van zulke zogenaamd ‘primitieve’ culturen en die verschillen onderling in allerlei opzichten. Met waterdieren hebben we opvallend veel kenmerken gemeen, en als we ooit volbloed jagers geweest zijn dan zijn roofdieren misschien ook wel een bruikbaar model. Een ‘objectieve’ keuze uit al die plausibele alternatieven is onmogelijk. Morris kiest roofdieren en moderne Amerikanen als model omdat hij daarin zijn eigen beeld van de mens het best gerepresenteerd ziet. Tanner kiest voor chimpansees en voor de zeer vredelievende bosjesmannen omdat ze zich daar het meest mee verwant voelt. Soortgelijke overwegingen gelden voor de andere auteurs.
De gedachte dat mannelijke biologen bij hun reconstructies van ons verre verleden hun eigen vooroordelen over de mens in hun verhalen hebben ingebouwd is niet nieuw. In zekere zin is dat de kern van de feministische kritiek op de verhalen van Morris en de zijnen en, recenter, op die van bijvoorbeeld Wilson en Barash. En die kritiek onderschrijf ik van harte. Alleen steekt het probleem net iets dieper. Ook de feministische theoretici zullen keuzes moeten doen. En ook zij zullen zich bij hun keuzes uiteindelijk laten leiden door hun mensbeeld. Er zit niets anders op. En dat betekent dat wat we uit een evolutionaire reconstructie kunnen leren over onze menselijke natuur nooit veel meer is dan wat de betreffende auteur daar zelf al van tevoren over dacht. Voor ik besluit dat dat voor evolutiebiologie in het algemeen geldt is het goed om nog even stil te staan bij de vraag of de evolutietheorie ons direct, zonder tussenkomst van mogelijk wankele reconstructies, iets over onze aard kan leren. Er zijn wetenschappers die dat lijken te denken.
5.2.3. evolutionaire karakterisering zonder reconstructie?
Sommige sociobiologen zijn kennelijk van mening dat je op basis van wat we nu aan theorie hebben met een minimum aan onomstreden feitenmateriaal al heel wat kunt zeggen over de aard en het gedrag van dieren. Een aardig voorbeeld in dit verband is de theorie van Trivers[213] over voortplanting en investering. Volgens hem zal de bereidheid van een individu om voor zijn of haar kind te zorgen afhangen van de hoeveelheid energie die dat individu in dat kind gestoken heeft. Die hoeveelheid is voor vrouwen in eerste instantie altijd groter dan voor mannen, een eitje kost nu eenmaal meer dan een paar zaadcellen. En bij, bijvoorbeeld, zoogdieren loopt het verschil ten gevolge van de draagtijd en de noodzaak om het kind te zogen enorm op. Wilson[214] meent dat dat op zich al voldoende grond biedt om een heleboel gedragsverschillen tussen mannen en vrouwen te verklaren. Vrouwen zijn relatief beperkt in het aantal kinderen dat ze kunnen krijgen. Het is dus in hun reproductieve voordeel om te zorgen dat de paar kinderen die ze krijgen welbeslagen ten ijs komen. Dat wil zeggen dat ze er goed aan doen de kat eens rustig uit de boom te kijken, niet met de eerste de beste man in zee te gaan, maar te wachten tot ze er een vinden die hen een stel gezonde, sterke en intelligente kinderen kan geven, en zo mogelijk een die bereid is zelf het nodige aan de opvoeding van zijn kroost bij te dragen. Mannen kunnen in theorie duizenden kinderen verwekken. Het zal hen dus minder deren als sommige daarvan geen succesvolle leden van de volgende generatie worden. Elke gelegenheid om een kind te verwekken zullen ze daarom enthousiast aangrijpen. Polygynie en een dubbele seksuele moraal zijn in dit licht eenvoudig te verklaren. Vrouwen zijn, als het om voortplanting gaat, de beperkende factor. De onderlinge concurrentie tussen mannen zal dus veel groter zijn dan die tussen vrouwen. Die concurrentie uit zich ook in een strijd tussen mannen onderling. Het is daarom begrijpelijk dat mannen groter, sterker en agressiever zijn dan vrouwen.
Over dit soort verklaringen is veel geschreven. Sommigen verwijten sociobiologen dat ze er een nu bestaande ongelijkheid tussen de seksen mee rechtvaardigen.[215] Dat verwijt is begrijpelijk, maar het richt zich ten onrechte op de theoretische uitgangspunten. Je kunt Wilson (en hem niet alleen) verwijten dat hij het verschil tussen verklaren en rechtvaardigen uit het oog verliest. Hij schrijft soms wel erg vergoelijkend over de verhouding tussen de seksen zoals die nu in onze samenleving lijkt te bestaan. Maar het heeft geen zin je onvrede met een bestaande situatie (even aangenomen dat je die situatie juist inschat) te richten op een theorie die die situatie verklaart.
Relevanter is de vraag of de theorie inderdaad een aanvaardbare verklaring van de bestaande situatie geeft. Daarvoor zouden we allereerst moeten weten of mannen inderdaad meer dan vrouwen neigen tot buitenechtelijke avonturen, of vrouwen zich meer dan mannen verplicht voelen al hun energie en aandacht te geven aan hun kinderen. En daarvoor kun je niet volstaan met de constatering dat zulks een gangbare opvatting is in onze laat twintigste-eeuwse westerse samenleving. We zouden meer moeten weten over het dagelijkse leven en streven van alle mensen in alle samenlevingen. Over die kennis beschikken sociobiologen niet. Het is niet onmogelijk dat mannen en vrouwen over het geheel genomen minder verschillen dan zij nu aannemen. Daaruit op zich zou overigens niet volgen dat de theorie waar sociobiologen mee werken ondeugdelijk is. Het betekent wel dat de feiten waar ze hun theorie op loslaten niet kloppen. En dat brengt me terug bij de hoofdlijn van mijn verhaal.
Het moge zo zijn dat mannen en vrouwen zich op karakteristieke manieren verschillend zullen gaan gedragen in situaties waarin ze verschillen wat betreft de hoeveelheid energie die ze in hun nageslacht investeren. Maar voor je zulke gedragsverschillen voorspelt zul je moeten vaststellen of aan je premissen is voldaan. Het is niet voldoende om vast te stellen dat eicellen zeldzamer en kostbaarder zijn dan zaadcellen. Beide geslachten investeren veel meer in hun nageslacht dan alleen hun geslachtscellen, en wel vóórdat er zelfs maar van nageslacht sprake is. Er gaat heel wat energie zitten in het zoeken en versieren van partners, het bouwen van nesten, het graven van holen, en het uit de buurt houden van andere kapers of kaapsters op de huwelijkskust. Voor je Trivers’ theorie over voortplanting en investering kunt toepassen zul je eerst een goed beeld moeten hebben van wie wanneer wat investeert. We zullen dus veel van mensen moeten weten voor we specifieke gedragsverschillen binnen onze soort, zo die er zijn, evolutionair kunnen verklaren. Een algemene theorie op zich is niet genoeg.
5.3. de menselijke natuur
Zoals we zagen komt een deel van de belangstelling voor ons evolutionaire verleden voort uit de gedachte dat we daaruit iets kunnen leren over hoe we echt zijn. En die kennis zou heel nuttig wezen. Gegeven de grote maatschappelijke en politieke problemen waar we voor staan, is het van belang om te weten wat we wel en wat we niet van mensen kunnen eisen en verwachten. Dat is dan ook precies de reden die bijvoorbeeld Wilson[216] geeft voor zijn speurtocht naar de menselijke natuur. En veel van de conclusies die hij trekt zijn in dienovereenkomstige termen vervat. We kunnen wel willen – de voorbeelden zijn van hem – dat mannen en vrouwen gelijke kansen krijgen op belangrijke posities in de samenleving, maar dan zullen we er rekening mee moeten houden dat mannen de wereld in willen om zich te bewijzen, terwijl vrouwen uiteindelijk liever thuis voor de kinderen zorgen. En voor de verhouding tussen mannen en vrouwen in de privésfeer gelden vergelijkbare overwegingen. Vrouwen zijn volgens Wilson monogamer ingesteld dan mannen, dat hangt samen met belangenverschillen waar het de voortplanting betreft. We kunnen van mensen wel willen dat ze zich net zo slaafs aan een taak zullen wijden als mieren en termietenwerkers, maar dat ligt niet in onze aard. Natuurlijk zijn wij heel plooibaar, maar die plooibaarheid heeft zijn grenzen. Er zijn fraaie idealen denkbaar die misschien alleen gerealiseerd zouden kunnen worden in een totalitaire staat. Kennis van de menselijke natuur is hoe dan ook onontbeerlijk voor wie zich bezig houdt met het ontwikkelen van politieke programma’s of ethische systemen. Tot zover Wilson. Ik denk niet dat wat hij beweert helemaal onzin is, maar er is wel reden voor voorzichtigheid en terughoudendheid. En daar wil ik dit boek mee besluiten.
Met ‘de menselijke natuur’ kun je verschillende dingen bedoelen. Sommige mensen zijn op zoek naar wat ons uniek maakt, naar wat ons onderscheidt van alle andere dieren: de menselijke natuur in enge zin. Anderen zijn al heel tevreden met een overzicht van wat mensen gemeenschappelijk hebben: de menselijke natuur in ruime zin. Laten we die twee eens een voor een bekijken.
5.3.1. de menselijke natuur in enge zin
Veel verhalen over de mens beginnen met een kenschetsing van de mens als bijzonder dier. En er zijn heel wat kenmerken die in zulke karakteriseringen worden opgevoerd. Ik noem er hier een paar, zonder bronvermelding.
De mens is een rationeel wezen, hij denkt, heeft besef van wat in de tijd voor en achter hem ligt, heeft weet van wat zich afspeelt op plaatsen waar hij niet is. De mens is een symbolisch en religieus wezen, hij heeft een taal, geeft betekenissen aan dingen en handelingen, creëert dingen en ontwikkelt handelingen soms louter vanwege de betekenis die ze voor hem hebben. De mens is een creatief, inventief en actief wezen, hij gebruikt gereedschappen, maakt met gereedschap nieuw gereedschap, hij cultiveert andere organismen en verandert ze naar het hem uitkomt. De mens is een moreel wezen, hij heeft een zelfbeeld, ontwikkelt zijn eigen ethiek en overschrijdt die soms ook. Hij voert oorlog, pleegt zelfmoord, doodt soortgenoten zonder ze te eten, eet soortgenoten. En voorzover de mens dan nog een beetje dier is is hij een bijzonder dier: naakt, staartloos en rechtopgaand, met relatief grote hersenen, een uitzonderlijk lange kindertijd en twee rechterhanden.
Ik zal niet al die facetten afzonderlijk bespreken. Maar een paar opmerkingen zijn van belang. Voor veel van deze kenmerken geldt dat we ze, bij nader inzien, bij andere dieren ook vinden: gereedschapgebruik, het cultiveren van andere dier- (en planten-) soorten. Voor andere geldt dat we in elk geval niet zeker weten dat ze bij andere dieren níet voorkomen: zelfbeelden, bijvoorbeeld, en opvattingen over wat hoort en niet hoort. Misschien ben ik overdreven voorzichtig. Natuurlijk weten we niet zeker dat een worm geen zelfbeeld heeft, maar erg voorstelbaar is het toch niet? Misschien is dat inderdaad moeilijk voorstelbaar, maar dat geldt voor wel meer zaken – de gekromde ruimtes van de relativiteitstheoretici, de antimaterie van de kernfysici – dus ons voorstellingsvermogen is een raadgever met beperkingen. Laten we ons oordeel als dat enigszins kan nog even opschorten.
Een ander probleem is serieuzer. De kenmerken die ik eerder opsomde zijn wel erg algemeen verwoord. De mens voert ongetwijfeld zo nu en dan oorlog, maar ik weet zeker dat de meesten van mijn lezers nooit in een oorlog hebben meegevochten, en dat ze dat allesbehalve betreuren. Dat is ook precies het probleem met uitspraken over ‘de’ mens: meestal weten we maar van een paar mensen zeker dat die uitspraken voor hen waar zijn. Als de menselijke natuur een uníek menselijke natuur moet zijn, zijn we gauw uitgepraat, en verder rest ons vooral onzekerheid.
5.3.2. de menselijke natuur in ruime zin
Wij vormen met z’n allen samen één soort. Betekent dat niet op zijn minst dat er een reeks van kenmerken moet zijn die we met zijn allen gemeen hebben en die we kunnen gebruiken om een beschrijving van de menselijk natuur in ruime zin te geven? Voor een antwoord op die vraag moeten we eerst iets beter kijken naar wat een soort precies is.
Soorten werden lange tijd beschouwd als groepen van individuen die een aantal anatomische en fysiologische eigenschappen met elkaar gemeen hebben. Maar er zijn nogal wat groepen die we intuïtief als soorten beschouwen waarvoor dat overduidelijk niet opgaat. Organismen waarbij generatiewisseling optreedt vormen daarvan de fraaiste illustratie. Het syphonale groenwier Halicystis[217] heeft een vertakte draadvormige geslachtelijke generatie en een ballonvormige ongeslachtelijke generatie. Die lijken zo weinig op elkaar dat ze lange tijd in verschillende soorten (en die telkens elk weer in een eigen geslacht: Halicystis en Derbesia) zijn ondergebracht. Nú worden de opeenvolgende generaties samen in één soort gestopt omdat biologen ‘biologische’ verwantschap (gedefinieerd in termen van afstammingsrelaties) een belangrijker criterium vinden dan vórmverwantschap. Die afweging sluit aan bij de overtuiging dat de leden van opeenvolgende generaties ten gevolge van geleidelijke genetische veranderingen vaak van elkaar zullen verschillen.
Zonder variatie in vorm en gedrag binnen een soort zou evolutie onmogelijk zijn. Voortbordurend op dit gegeven kwamen Ghiselin en Hull[218] tot de suggestie dat we een soort[219] niet moeten opvatten als een verzameling van individuele organismen maar als een individu waarvan de verschillende individuele organismen evenzovele onderdelen vormen. Dat grote individu verandert in de loop van de tijd. En net zo goed als de onderdelen van een individueel organisme van elkaar verschillen zullen de leden – liever dus: onderdelen – van een soort van elkaar verschillen. Over de vraag of het zin heeft een soort als een individu op te vatten valt te twisten,[220] maar de basis van die opvatting, het inzicht dat de leden van een soort van elkaar verschillen, blijft daarbij overeind. En voor een discussie over de menselijke natuur is dat een relevante gedachte. Evolutionaire overwegingen leiden tot de conclusie dat soorten geen ‘natuur’ hoeven te hebben als je de natuur van een soort opvat als een reeks eigenschappen die alle leden van die soort gemeen hebben. We zullen met generaliseringen heel voorzichtig moeten zijn.
Dat werpt een ander licht op het soort vragen waar we even terug mee werden geconfronteerd. Zijn vrouwen monogamer ingesteld dan mannen? Hebben vrouwen een onontkoombaar moederinstinct? Als soorten inderdaad zo variabel zijn als evolutiebiologen (moeten) aannemen, dan ligt een antwoord als ‘sommigen wel, anderen niet’ het meest voor de hand.
Op deze conclusie valt misschien wel wat af te dingen. Volgens sommige ethici is er bijvoorbeeld duidelijk sprake van moréle universalia, van ethische principes die we in alle culturen terugvinden. En sociobiologen die door ethiek worden gefascineerd werpen zich met graagte op deze universele beginselen.
5.4. morele universalia
In alle menselijke samenlevingen zijn moord en doodslag, diefstal, incest en verkrachting, verboden, en bijna overal worden hulpvaardigheid, eerlijkheid, en zorgzaamheid ten opzichte van kinderen en ouden van dagen, als deugden gezien. Dat doet vermoeden dat mensen als het op hun morele oordelen aankomt heel wat overeenkomsten vertonen. Maar soms bedriegt de schijn.
We zagen in hoofdstuk 4 dat mensen heel verschillende dingen kunnen bedoelen als ze een bepaalde handeling op morele gronden veroordelen. Twee mensen van wie de een het slecht vindt om een medemens van het leven te beroven omdat dat het gemiddelde geluk nadelig zou beïnvloeden, terwijl de ander hetzelfde vindt omdat de mens naar Gods beeld geschapen is en het daarmee overeenstemmende respect verdient, zullen zich doorgaans dezelfde beperkingen tegenover andermans leven opleggen (even afgezien van mogelijke temperamentsverschillen). Maar er zijn situaties denkbaar waarin de eerste het doden van een medemens goed zou vinden, bijvoorbeeld omdat dat de dood van onschuldige soortgenoten voorkomt, terwijl de tweede zulk ingrijpen zou afkeuren.[221] Binnen een samenleving vertonen mensen vaak grote overeenkomsten in gedrag, maar dat betekent niet dat er geen relevante verschillen zijn. Alleen, we krijgen die niet altijd even snel te zien. We zijn soms geneigd verschillen in gedrag in ongewone, spanningsvolle, situaties toe te schrijven aan verschillen in zelfbeheersing. Ik denk dat we daar voorzichtig mee moeten zijn. We redden ons geloof in morele universalia, maar we negeren de mogelijkheid van verschillen in morele uitgangspunten.
Mijn twijfel aan het bestaan van universele morele overtuigingen wordt versterkt door wat ik lees over verschillen tussen menselijke samenlevingen. Het moge zo zijn dat moord overal als afkeurenswaardig wordt gezien, maar niet overal bedoelt men met moord hetzelfde. Het is nog niet zo lang geleden dat ook in sommige kringen in onze eigen samenleving een ernstige belediging als een goede reden werd beschouwd om een eerlijk duel aan te gaan. En ook nu nog zijn er samenlevingen waarin bloedwraak als een kwestie van eer wordt beschouwd. Iedereen is tegen moord, maar niet iedereen geeft hetzelfde antwoord op de vraag wanneer er van ‘moord’ sprake is. Iedereen is tegen verkrachting, maar de heftige discussies over de vraag of er binnen het huwelijk van verkrachting sprake kan zijn doen vermoeden dat we het niet eens zijn over de vraag wat ‘verkrachting’ betekent. Voor incest geldt hetzelfde. Het is overal verboden, maar er zijn samenlevingen waarin een neef of nicht, mits van de juiste zijde van de familie, als de meest ideale huwelijkspartner wordt beschouwd,[222] terwijl men in andere samenlevingen al bij een huwelijk met een zwager of schoonzus van incest begint te spreken.
Voor andere morele principes geldt hetzelfde. Edel en Edel beschrijven het verschil in opvatting over eerlijkheid en de plicht om aangegane verplichtingen na te leven tussen de bewoners van twee naburige eilandjes in de Stille Oceaan. Op het ene eiland zullen bewoners zich alle moeite getroosten om een handelspartner een wederdienst te bewijzen, zelfs als ze daardoor een kans om goed geld te maken verspelen, terwijl de bewoners van een naburig eiland leugen en bedrog als een blijk van grote levenskunst beschouwen.[223]
Je zou je kunnen afvragen of de schijnbare overeenstemming over morele beginselen tussen verschillende samenlevingen niet vooral het gevolg is van een vertaalfout. Ik weet niet welk woord de oorspronkelijke Australiërs (‘aborigines’) gebruiken als ze een huwelijk met een bepaalde partner veroordelen vanwege de aard en de mate van verwantschap, maar ik vermoed dat het misleidend is dat woord, zoals men meestal doet, met ‘incest’ te vertalen. Die vertaling verhult een verschil in inzicht en draagt dus niet bij tot wederzijds begrip. Het enige dat wellicht echt universeel is, is ’s mensen neiging zich te houden aan een stelsel van regels. Maar dat is geen inzicht waar het ontwerpen van een morele code heel veel makkelijker door wordt.
5.5. sociobiologie en morele universalia
Veel sociobiologen geloven niet alleen in het bestaan van morele universalia, ze menen ook dat ze die kunnen verklaren. Zou dat ons vertrouwen in het bestaan van een universele morele menselijke natuur niet weer een beetje kunnen herstellen? Ik denk het niet. Ik zal die twijfel aan de hand van een voorbeeld toelichten.
Over incest is door sociobiologen veel geschreven.[224] Het biedt daarom een goed voorbeeld van de verheldering die we van de sociobiologie kunnen verwachten. Uitgangspunt van de sociobiologische verklaring is dat incest een genetisch risico met zich brengt. Een kind geboren uit een incestueuze relatie heeft een grotere kans op homozygotie. Dat wil zeggen dat genen op dezelfde plek op bij elkaar horende chromosomen gelijk zijn. En homozygotie is een risico in geval van een recessief gen dat tot minder wenselijke kenmerken leidt. Zo’n gen (preciezer: allel) zal als het niet te algemeen is zelden tot expressie komen omdat het meestal als buur een dominant gen (allel) met minder kwalijke gevolgen heeft. Het zal dus niet snel door selectie verdwijnen. Gedrag dat tot incestvermijding leidt is daarom functioneel. Het komt dan ook bij zeer veel soorten voor. Het simpelste mechanisme is een verspreiding van individuen vóór de geslachtsrijpe leeftijd, waardoor de kans dat broertjes en zusjes elkaar tegenkomen als ze naar een partner uitzien klein wordt. Maar bij soorten waar kinderen tot vlak voor of zelfs na hun geslachtsrijpheid door hun ouders worden opgevoed kan dat natuurlijk niet. Mensen vormen een goed voorbeeld van zo’n soort. In zulke gevallen moeten andere mechanismen het werk overnemen. En een van de dingen die bij ons, volgens veel sociobiologen, die functie vervult is de milde en onbewuste afkeer die mensen hebben van seks met de leeftijdsgenoten met wie ze opgroeiden. Broertjes en zusjes worden gewoon niet verliefd op elkaar, zelfs niet als ze verstandelijk heel wel inzien dat de ander een aantrekkelijke huwelijkspartner is.[225] Het incesttaboe zou van die, aangeboren, afkeer de maatschappelijke vertaling zijn.
Er kleven een aantal bezwaren aan deze verklaring. Als de neiging om incest te vermijden zo diep verankerd is als de verklaring veronderstelt is het onduidelijk waarom er nog van uitgebreide morele verhandelingen en wetgeving met betrekking tot incest sprake zou moeten zijn. Bovendien komt incest wel degelijk op ruime schaal voor. Voorzover er sprake is van een taboe lijkt het soms eerder taboe om over incest te spreken, dan om het te bedrijven. En er zijn, zoals we zagen, samenlevingen waarin iets dat velen van ons als incest zouden beschouwen, een huwelijk tussen derdegraads verwanten, wordt gepropageerd.
Daar zou de rechtgeaarde sociobioloog tegen in kunnen brengen dat het mechanisme niet waterdicht is, en dat we dat ook niet mogen verwachten. Het is ontstaan in een periode waarin het leven van mensen er anders uitzag dat het onze nu. Mensen hebben (als we het begrip ‘mens’ niet al te strikt nemen) miljoenen jaren geleefd in kleine groepen met veel sociale controle en weinig privacy. Ze moesten hard werken, werden niet erg oud, en trouwden (neem dat woord niet te letterlijk) al jong. De schrijnende incestgevallen waar we nu regelmatig mee worden geconfronteerd komen in zo’n situatie wellicht veel minder voor. En het blijft zo dat er ook nú maar weinig kinderen worden geboren van wie de ouders in de eerste of tweede graad verwant zijn. En daar heeft die sociobioloog natuurlijk gelijk in. Mensen gedragen zich in elk geval meestal zo dat hun kinderen ouders krijgen die niet al te nauw verwant zijn. En dat is gezien de risico’s verbonden aan een te nauwe verwantschap tussen ouders, ook wel begrijpelijk.[226] Maar het is de moeite waard om preciezer te formuleren wat er vermeden wordt. En daarbij is het handig om onderscheid te maken tussen ‘incest’ en ‘inteelt’.
‘Inteelt’ is een puur biologisch begrip, gedefinieerd in termen van de mate van genetische verwantschap, en het heeft direct betrekking op voortplanting. We spreken van inteelt als er nageslacht geboren wordt uit een relatie tussen individuen die genetisch gezien nauw verwant zijn. De ‘incest’ waar antropologen en ethici het over hebben is iets anders. Daarbij gaat het om seksuele contacten tussen individuen die een relatie met elkaar hebben waarbinnen seksueel contact door de samenleving waarvan zij deel uitmaken, niet wordt toegestaan omdat ze elkaar te na staan. Zo heet seksueel contact tussen pleegzus en pleegbroer in onze samenleving incest, zelfs als dat contact niet zo ver gaat dat het tot zwangerschap kan leiden. Verder is inteelt een relatief begrip. Een voortplantingsrelatie tussen broer en zus is meer inteelt dan een dergelijke relatie tussen neef en nicht. Wat de bioloog verklaren kan is het vermijden van een te grote mate van inteelt. Over incest zegt dat maar zeer ten dele iets. Dus een biologische verklaring van ‘incest’-vermijding leert ons weinig met betrekking tot morele oordelen over incest in de dagelijkse zin van dat woord (en daar gáán morele oordelen over incest over!).
Sociobiologen zijn vast ook in staat om onze bezwaren tegen moord en doodslag, diefstal en verkrachting te verklaren. Maar daarbij zullen zich dezelfde problemen voordoen. Sociobiologische verklaringen laten de grote variatie aan oordelen binnen en tussen samenlevingen onverlet. Daarmee zeg ik niet dat die verklaringen overal en altijd irrelevant zijn. Een beetje begrip is ook begrip. Maar wie geïnteresseerd is in mensen en hun bonte verscheidenheid heeft aan evolutiebiologie niet zo veel.
besluit
Geen enkel onderzoek is ooit af. Elke conclusie is voorlopig, en er blijven altijd dingen over die nader onderzoek behoeven. Als het een beetje wil, zijn er in de loop van een onderzoek zelfs van zulke dingen bijgekomen! Zo bezien is er alle reden om het verslag van een onderzoek gewoon maar, liefst midden in een zin, te laten ophouden. Maar dat is zo onbevredigend. Daarom besluit ik mijn verhaal met een (tussentijdse) balans.
Elke discussie over sociobiologie wordt verstoord door over-en-weerse tegenwerpingen in termen van één grote tweedeling, met natuurlijke, genetisch bepaalde, aangeboren, biologische, kenmerken aan de ene, de dierlijke, kant, en culturele, omgevingsbepaalde, aangeleerde, sociaal bepaalde kenmerken aan de anderen, menselijke, kant (zie afb. 2.1). Het heeft dus zin om eerst en vooral af te rekenen met dat dichotome denken. Hoe overzichtelijk de grote tweedeling ook zijn moge, zij is uiterst misleidend. Dat betekent niet dat de verschillende begrippen uit die tweedeling nu verder taboe zijn, maar in elke geval wel dat ze niet in twee grote groepen tegenover elkaar gezet moeten worden.
Ik denk verder dat (evolutie)biologen er goed aan zullen doen het woord ‘cultuur’ te mijden. Alles wat ze daar op dit moment mee kunnen bedoelen wordt prima gedekt door een begrip als ‘omgevingsinvloeden’. Dat geeft dan meteen aan waar de grote uitdaging voor de evolutiebiologie ligt: het ontwikkelen van een theorie waarin de invloeden van zowel genetische als omgevingsveranderingen op de verschijningsvormen van het leven op aarde beschreven worden.
Het bestaan van altruïstisch gedrag werd door veel sociobiologen beschouwd als een groot probleem, omdat dat in strijd leek met gangbare theorieën uit de evolutiebiologie. Ze hebben voor dat probleem ingenieuze oplossingen bedacht, maar het ziet ernaar uit dat ze daarbij een belangrijke stap hebben overgeslagen. Een analyse van het gebruikte begripsapparaat suggereert dat het altruïsmeraadsel op zich een schijnprobleem is, dat ontstond door begripsverwarring. Dat betekent natuurlijk niet dat sociobiologisch onderzoek aan hulpgedrag bij allerlei dieren net zo goed niet gedaan had kunnen worden. Dat onderzoek heeft onze kennis van hoe dieren met elkaar omgaan vergroot.
De gedachte dat sociobiologen iets zinnigs zouden kunnen zeggen over altruïstisch gedrag vormt voor velen hunner een eerste reden om te denken dat sociobiologische theorieën ethische relevantie hebben. Bij die gedachte passen enige kanttekeningen. Bioaltruïsme in een ethologische zin des woords kan met enige moeite wel met ethisch altruïsme in verband worden gebracht. Dat leidt in principe tot de mogelijkheid (sommige) vormen van (ethisch) altruïsme te verklaren en inzicht te verwerven in de omstandigheden waaronder dat gedrag zich zal voordoen. Bioaltruïsme in een evolutionair relevante zin des woords heeft, zowel door een verschil in logische structuur als door een verschil wat betreft de grootheid waarin verlies en winst worden berekend, geen direct verband met altruïsme in een ethologische zin, en al helemaal niet met altruïsme in een ethische zin des woords. Evolutionaire overwegingen hebben daarom weinig belang voor ons inzicht in de mogelijkheid van ethisch altruïsme. Laat staan in de wenselijkheid daarvan.
Ook anderszins is er weinig reden om in ethische discussies erg veel rekening te houden met evolutionaire overwegingen. Elke poging om ethische principes te grondvesten in evolutietheorie loopt spaak. En zelfs een afstandelijker verband tussen evolutietheorie en ethiek is beduidend minder solide dan sommige sociobiologen denken. Uitspraken over de menselijke natuur die een rol kunnen spelen in ethische discussies laten zich uit de resultaten van evolutionair onderzoek niet afleiden. Het is zelfs de vraag of het zin heeft om te spreken over zoiets als een menselijke natuur.
De discussie over de relatie tussen evolutiebiologie en ethiek laaide flink op toen Wilson voorstelde de ethiek te verbiologiseren. Ik hoop dat ik heb duidelijk gemaakt waarom dat voorstel niet moet worden aangenomen. Maar ik moet ervoor oppassen niet in een ander uiterste te vervallen. Alles wat tot mensenkennis leidt is van belang voor wie nadenkt over morele beginselen en hun praktische consequenties. Ook biologisch, en dan met name ethologisch, onderzoek aan mensen leidt tot mensenkennis. Ik ben vrij sceptisch ten aanzien van het relatieve belang van dat onderzoek, maar dat is ten dele een gevoelskwestie, en niet een onontkoombare conclusie uit mijn eigen onderzoek.
Veel van de conclusies uit mijn onderzoek zijn het makkelijkst in de vorm van ontkenningen te formuleren: dit moet niet, dat mag niet, dit kan niet, dat werkt niet. Op sommige lezers zal dat een negatieve indruk maken, maar het schept ruimte voor wat er wel mag, kan en moet. Een paar woorden daarover.
In de eerste plaats zou er meer onderzoek moeten worden gedaan aan de invloed van omgevingsvariatie op fenotypische variatie. Dit alleen al om te voorkomen dat we ons blind staren op de rol van genetische variatie. Dat is niet alleen van belang voor de sociobiologie, maar ook bijvoorbeeld voor discussies over de mogelijkheden en beperkingen van genetische manipulatie.
Vervolgens zouden sociobiologen (en evolutiebiologen in het algemeen) moeten proberen een theorie te ontwikkelen over het verband tussen genetische veranderingen, omgevingsveranderingen, en fenotypische veranderingen van generatie op generatie.
Ik liet hierboven de mogelijkheid open dat ethologisch onderzoek ons meer inzicht zou kunnen geven in vormen van sociaal gedrag die verband houden met wat we moreel relevant achten. Voor we een definitief oordeel over het belang van zulke kennis kunnen geven zal er veel meer onderzoek in deze richting moeten worden gedaan.
Tenslotte denk ik dat er meer aandacht zou moeten worden besteed aan het soort analyses waarvan dit boekje voorbeelden bevat. Wetenschap zonder begripsmatige helderheid raakt verstrikt in schijnproblemen. En dat niet alleen: het feit dat velen gefascineerd raken door de belofte van een diepgaand inzicht in ons eigen lot, ons door de sociobiologie geboden, laat zien dat onheldere ‘wetenschap’ een zelfstandige maatschappelijke kracht kan worden. We doen er goed aan die kracht te breken. De erosie waaraan traditionele wereldbeelden, met name die van de grote religies, op dit moment onderhevig zijn, schept een vacuüm waarbinnen nieuwe ideologieën een kans krijgen. Ik denk dat het niet goed is wanneer wetenschap de belangrijkste leverancier van zulke ideologieën wordt. Dat geldt zelfs voor wetenschap op zijn best. Wetenschap moet (per definitie!) afstandelijk zijn. Dat levert boeiende kennis op, maar wijsheid garandeert het niet.
Newton zag zichzelf als een kind dat aan het zeestrand speelde en zo nu en dan een gladdere steen, een mooiere schelp ontdekte dan anderen, terwijl de grote oceaan der wereld onontdekt voor hem lag. Misschien is onze schelpenverzameling wat groter dan de zijne, maar van de zee weet wetenschap nog altijd niets.
geciteerde literatuur
Allen, E., c.s., 1978. Against ‘sociobiology’. In: Caplan 1978: 259-264.
Banning, W., 1972. Typen van zedenleer. Bohn, Haarlem.
Barash, D.P., 1979. Sociobiology: the whisperings within. Harper & Row, New York.
Barker, M., 1981. The new racism – conservatives and the ideology of the tribe. Junction books, Londen.
Barlow, G.W. en J. Silverberg (red.), 1980. Sociobiology: beyond nature/nurture? Westview Press, Boulder, Colorado.
Barnett, S.A., 1980. Biological determinism and the Tasmanian native hen. In: Montagu 1980: 135-157.
Beatty, J., 1984. Chance and natural selection. Philosophy of Science 51: 183-211.
Berghe, L.P. van den, 1980. Incest and exogamy: a sociobiological reconsideration. Ethology and Sociobiology 1: 151-162.
Bertram, B.C.R., 1982. Problems with altruism. In: Kings College Sociobiology Group 1982: 251-267.
Bonner, J.T., 1980. The evolution of culture in animals. Princeton up, New Jersey.
Boon, L. en H. Smit, 1985. Sociobiologische onderzoeksstijl en popularisering. Kennis en Methode ix (4): 319-333.
Boorman, S.A. en P.R. Levitt, 1980. The genetics of altruism. Academic Press, New York.
Boyd, R. en P.J. Richerson, 1985. Culture and the evolutionary process. University of Chicago Press, Chicago.
Campbell, D.T., 1978. Social morality norms as evidence of conflict between biological human nature and social system requirements. In: Stent 1978: 67-82.
Caplan, A.L., 1978. The sociobiology debate, readings on ethical and scientific issues. Harper & Row, New York.
Caplan, A.L., 1980. A critical evaluation of current sociobiological theory: adequacy and implications. In: Barlow en Silverberg 1980: 97-113.
Cavalli-Sforza, L.L. en M.W. Feldman, 1981. Cultural transmission and evolution: a quantitative approach. Princeton up, Princeton.
Churchland, P.S., 1986. Neurophilosophy, towards a unified science of the mind-brain. mit Press, Cambridge, Massachusetts.
Clutton-Brock, T.H. en P.H. Harvey (red.), 1978. Readings in sociobiology. Freeman, Reading.
Darwin, C., 1871. The descent of man and selection in relation to sex. Heruitgegeven door Random House, New York (zonder jaartal).
Dawkins, R. 1976. The selfish gene. Oxford up, Oxford.
Dawkins, R. 1979. Twelve misunderstandings concerning kin selection. Zeitschrft für Tierpsychologie 51: 184-200.
Dawkins, R. 1982. The extended phenotype. Freeman, Oxford.
Denffer, D. von, W. Schumacher, K, Magdefrau en F. Firbas, 1967. Lehrbuch der Botanik für Hochschule (‘Strasburger’). Fischer, Stuttgart.
Dobzhansky, T.G. en E. Boesiger, 1983. Human culture: a moment in evolution. Columbia up, New York (uitgave verzorgd door B. Wallace).
Dominey, W.J., 1984. Alternative mating tactics and evolutionary stable strategies. American Zoologist 24: 385-396.
Edel, M. en A. Edel, 1968. Anthropology and ethics – the quest for moral understanding. The Press of Case Western Reserve University, Cleveland.
Edwards, P.(eindred.), 1967. The encyclopedia of philosophy. Macmillan & The Free Press, New York.
Faber, J., 1986. Holisme als noodzakelijke benaderingswijze in de embryologie. Vakblad voor Biologen 66: 241-243.
Falconer, D.S., 1981. Introduction to quantitative genetics. (Tweede druk.) Longman, Londen.
Foot, P., 1967. Theories of ethics. Oxford University Press, Oxford.
Fossey, D., 1984. Gorilla’s in de mist. (Vertaling: H. Heij.) Veen, Utrecht.
Freeman, D., 1980. Sociobiology: the ‘antidiscipline’ of anthropology. In: Montagu 1980: 198-219.
Ghiselin, M.T., 1966. On psychologism in the logic of taxonomic controversies. Systematic Zoology 15: 207-215.
Gould, S.J., 1980. Sociobiology and human nature: a postpanglossian vision. In: Montagu 1980: 283-290.
Graaf, J. de, 1972. Elementair begrip van de ethiek. Bohn, Haarlem.
Gupta, A.P. en R.C. Lewontin, 1982. A study of reaction norms in natural populations of Drosophila pseudoobscura. Evolution 36: 934-948.
Hamilton, W.D., 1964. The genetic evolution of social behavior. Journal of Theoretical Biology 7: 1-6.
Hartog, H. den, 1981. Sociobiologie: een oude hypothese verpakt als nieuwe synthese. In: De Waal 1981: 133-151.
Heeger, R., 1985. Utilisme en aanvaardbaarheid. In: Hubbeling en Veldhuis 1985: 18-47.
Hooff, J.A.R.A.M. van, 1981. Inleiding in de evolutionaire verklaring van sociaal gedrag. In: De Waal 1981: 7-36.
Houtman, C., c.s., 1986. Schepping en evolutie. Kok, Kampen.
Hubbeling, H.G. en R. Veldhuis (red.), 1985. Ethiek in meervoud. Van Gorkum, Assen.
Hudson, W.D., 1983. Modern moral philosophy. (Tweede druk.) Macmillan, Londen.
Huis in ‘t Veld, G., 1982. Beestachtige sprookjes, de sociobiologie brengt dubbel onheil. Intermediair 18(26): 25-29.
Hull, D.L., 1974. Philosophy of biological science. Prentice Hall, Englewood Clifs.
Hull, D.L., 1978. A matter of individuality. Philosophy of Science 45:335-360.
Hull, D.L., 1980. Individuality and selection. Annual Review of Ecology and Systematics 11: 311-332.
Kalsbeek, L., 1970. De wijsbegeerte der wetsidee – proeve van een christelijke filosofie. Buijten en Schipperheijn, Amsterdam.
King, J.C., 1980. The genetics of sociobiology. In: Montagu 1980: 82-107.
King’s College Sociobiology Group (red.), 1982. Current problems in sociobiology. Cambridge up, Cambridge.
Kitcher, P., 1982. Abusing science, the case against creationism. mit Press, Cambridge, Massachusetts.
Kitcher, P., 1985. Vaulting ambition, sociobiology and the quest for human nature. mit Press, Cambridge, Massachusetts.
Kroeber, A.L. en C. Kluckhohn, 1952. Culture: a critical examination of concepts and definitions. Random House, New York.
Kurland, J.A., 1977. Kin selection in the Japanese monkey. Contributions to Primatology 12: 1-145.
Leaky, R.E. en R. Lewin, 1978. People of the lake – mankind and its beginnings. Avon Books, New York.
Lehrman, D.S., 1970. Semantic and conceptual issues in the nature- nurture problem. In: L.R. Aronson (red.): Development and evolution of behavior – essays in memory of T.C. Schneirla: 17-52. Freeman, San Francisco.
Lever, J. 1966. Dc interne ‘relatie’ van natuur en cultuur. Wijsgerig Perspectief op Maatschappij en Wetenschap 5(5): 257-275.
Lewontin, R.C., 1970. The units of selection. Annual Review of Ecology and Systematics 1:1-18.
Lewontin, R.C., 1974. The genetic basis of evolutionary change. Colombia up, New York
Lopreato, J., 1984. Human nature and biocultural evolution. Allen & Unwin, Boston.
Lorenz, K., 1966. On agression. Methuen, Londen.
Lumsden, C.J. en E.O. Wilson, 1981. Genes, mind, and culture. Harvard up, Cambridge, Massachusetts.
Lumsden, C.J. en E.O. Wilson, 1982. Précis of genes, mind, and culture, and open peer commentary. Behavioral and Brain Sciences 5: 1-37.
Lumsden, C.J. en E.O. Wilson, 1983. Promethean fire – reflections on the origin of mind. Harvard up, Cambridge, Massachusetts.
Mackie, J.L., 1977. Ethics, inventing right and wrong. Penguin books, Harmondsworth.
Maris. C.W., 1981. Critique of the empiricist explanation of morality. Kluwer, Deventer.
Maxwell, M., 1984. Human evolution – a philosophical anthropology. Croom Helm, Londen.
Maynard Smith, J., l982a. Evolution and the theory of games. Cambridge up, Cambridge, Massachusetts.
Maynard Smith, J., l982b. Mind, and the linkage between genes and culture. Behavioral and Brain Sciences 5: 20-21.
McAlester, A.L., 1968. The history of life. Prentice Hall, Englewood Cliffs.
Meulenbelt, A., 1984. De schillen van de ui. sara, Amsterdam.
Montagu, A. (red.), 1980. Sociobiology examined. Oxford up, Oxford.
Moore, G.E., 1942. A reply to my critics. In Foot 1967: 33-49.
Morgan, E., 1972. The descent of woman. Bantam Books, New York.
Morgan, E., 1982. The aquatic ape, a theory of human evolution. Souvenir Press, Londen
Morris, D., 1967. De naakte aap. (Vertaling T. Nicolaas.) Bruna, Utrecht.
Murphy, J.G., 1982. Evolution, morality, and the meaning of life. Rowman & Littlefield. Totowa.
Musschenga, A.W., 1981. De bijdrage van de sociobiologie aan moraalwetenschap en ethiek. In: De Waal 1981: 157-174.
Musschenga, A.W. (red.), 1984. Onderwijs in de natuurwetenschappen en morele vorming. Ten Have, Baarn.
Noske, B., 1984. Zijn dieren groen? Krisis 16: 5-15.
Richards, R.J., 1986a. A defense of evolutionary ethics. Biology and Philosophy 1: 265-293.
Richards, R.J., 1986b. Justification through biological faith: a rejoinder. Biology and Philosophy 1: 337-354.
Rose, S., 1980. ‘It’s only human nature’: the sociobiologist’s fairyland. In Montagu 1980: 158-170.
Roughgarden, J., 1979. Theory of population genetics and evolutionary ecology: an introduction. Macmillan, New York.
Ruse, M. 1973. The philosophy of biology. Hutchinson, Londen.
Ruse, M. 1979. Sociobiology: sense or nonsense? Reidel, Dordrecht.
Ruse, M. 1982. Darwinism defended – a guide to the evolutionary controversies. AddisonWesley, Londen.
Ruse, M. 1984a. The morality of the gene. The Monist 67: 167-199.
Ruse, M. 1984b. Is there a limit to our knowledge of evolution? BioScience 34: 100-104.
Ruse, M. en E.O. Wilson, 1985. The evolution of ethics. New Scientist, oktober 1985: 50-52.
Sahlins, M., 1977. The use and abuse of biology. Tavistock, Londen.
Sayers, J., 1982. Biological politics – feminist and anti-feminist perspectives. Tavistock, Londen.
Schellekens, H., 1985. Bouwstenen van de erfelijkheid – toepassing van recombinant DNA technologie in de geneeskunde. Meulenhof Informatief, Amsterdam.
Schlegel, H.G., 1969. Allgemeine Mikrobiologie. Thieme, Stutgart.
Schuyt, C.J.M., 1981. Sociobiologie en sociale wetenschappen: een kritische bijdrage. In: De Waal 1981: 200-204.
Simon, M.A., 1980. Biology, sociobiology, and the understanding of human social behavior. In: Montagu 1980: 291-310.
Singer, P., 1981. The expanding circle. ethics and sociobiology. Clarendon Press, Oxford.
Sloep, P.B., 1983. Patronen in het denken over vegetaties: een kritische beschouwing over de relatietheorie. Proefschrift, ru Groningen.
Sloep, P.B., 1986. Methodologie, waarom zou men zich er iets van aantrekken? Vakblad voor Biologen 66: 401-404.
Sober, E., 1984. The nature of selection: evolutionary theory in philosophical focus. mit Press, Cambridge, Massachusetts.
Sober, E. en R.C. Lewontin. 1982. Artifact, cause and genic selection. Philosophy of Science 49: 157-180.
Stebbins, G.L., 1982. Darwin to DNA, molecules to humanity. Freeman, San Francisco.
Steen, W.J. van der, 1982. Algemene methodologie voor biologen. Bohn, Scheltema en Holkema, Utrecht.
Steen, W.J. van der, 1984a. Grenzen van de wetenschap. In Musschenga 1984: 26-37.
Steen, W.J. van der, l984b. Methodological aspects of migration and orientation in fishes. In: J.D. McCleave (red.), 1984. Mechanisms of migration in fishes, 412-444. Plenum, New York.
Steen, W.J. van der, en B. Voorzanger, 1984a. Methodological problems in evolutionary biology iii. Selection and levels of organization. Acta Biotheoretica 33: 199-213.
Steen, W.J. van der, en B. Voorzanger, 1984b. Sociobiology in perspective. Journal of Human Evolution 13: 25-32.
Steen, W.J. van der, en B. Voorzanger, 1986. Methodological problems in evolutionary biology vii. The species plague. Acta Biotheoretica 35: 205-221.
Steen, W.J. van der, en P.J. Thung, 1988. Philosophy of the life sciences I, The coherence of medicine. Nijhoff, Dordrecht.
Stent, G.S. (red.), 1978. Morality as a biological phenomenon. University of California Press, Berkeley.
Tanner, N.M., 1981. On becoming human. Cambridge up, Cambridge.
Thomas, L., l986. Biological moralism. Biology and Philosophy 1: 316-325.
Trigg, R., 1986. Evolutionary ethics. Biology and Philosophy 1: 325-336.
Trivers, R.L., 1971. The evolution of reciprocal altruism. Quarterly Review of Biology 46: 35-47.
Trivers, R.L., 1974. Parent-offspring conflicts. American Zoologist 14: 249-264.
Trivers, R.L., 1985. Social evolution. Benjamin/Cummins, Menlo Park.
Toates, F., 1986. Motivational systems. Cambridge up, Cambridge.
Uyenoyama, M. en M.W. Feldman. 1980. Theories of kin and group selection: a population genetics perspective. Theoretical Population Biology 17(3): 380-414.
Voorzanger, B., 1982. Sociobiologie van de mens. lntermediair 18(30): 33.
Voorzanger, B., 1983. Eigenbelang en goede bedoelingen. Vakblad voor Biologen 63: 102-104.
Voorzanger, B., 1984a. Het mensbeeld in de biologie. In Musschenga 1984: 115-130.
Voorzanger, B., 1984b. Altruism in sociobiology, a conceptual analysis. Journal of Human Evolution 13:33-39.
Voorzanger, B. en P.B. Sloep, 1986. Boon en Smits populaire onderzoeksstijl. Kennis en Methode X(4): 361-366.
Voorzanger, B., 1987a. Methodological problems in evolutionary biology viii. Biology and culture. Acta Biotheoretica 36:23-34.
Voorzanger, B., 1987b. No norms and no nature – the moral relevance of evolutionary biology. Biology and Philosophy 2:253-270.
Vroon, P.A., 1981. Problemen? Voor alles is gewoon een gen. In: De Waal 1981: 99-118.
Waal, F.B.M. de (red.), 1981. Sociobiologie ter discussie. Bohn, Scheltema en Holkema, Utrecht.
Washburn, S.L., 1980. Human behavior and the behavior of other animals. In: Montagu 1980: 254-282.
Wilson, E.O., 1975. Sociobiology, the new synthesis. Harvard up, Harvard, Massachusetts.
Wilson, E.O., 1978. On human nature. Harvard up, Harvard, Massachusetts.
Wynne-Edwards, V.C., 1963. Intergroup selection in the evolution of social systems. Herdrukt in: Clutton-Brock en Harvey 1978: 10-19.
Zihlman, A. 1984. Pygmy chimps, people and pundits. New Scientist, 15 november 1984: 39-40.
Ik beloofde in een noot op blz. 105 nog een toevoeging over ‘fitness’ (daar weggelaten omdat lange voetnoten een fatsoenlijke opmaak onmogelijk maken):
Eén (probleem met ‘fitness‘) noem ik hier om de lezer in elk geval een beeld te geven van het curieuze karakter van dat begrip. Er zijn allerlei soorten waarvan de leden doodgaan voor ze nageslacht hebben. Ze hebben hun geslachtscellen geloosd, maar van een feitelijke bevruchting van eicellen is nog geen sprake. En andersom geldt voor heel wat organismen dat hun kindertal na hun dood nog behoorlijk kan dalen. Dus als je fitness wilt definiëren in termen van aantallen kinderen aan het eind van de levensweg moet je dat ‘eind’ niet al te letterlijk nemen. Hier valt dus nog wel wat te verfijnen. En daar komt nog iets bij. Fitness wordt vaak gedefinieerd niet in termen van aantallen kinderen zonder meer, maar van aantallen vruchtbare kinderen. Daar steekt een zeer redelijke gedachte achter. Wil fitness een evolutionair relevant begrip zijn dan zal het iets te maken moeten hebben met aantallen nakomelingen op de lange termijn. En wie zeer veel kinderen krijgt die allemaal steriel zijn hééft op de lange termijn geen nageslacht. (In zekere zin is niet alleen het al dan niet vruchtbaar zijn, maar ook de mate van vruchtbaarheid van kinderen van belang. Wie maar één kind krijgt dat voorbestemd is om heel veel kinderen te krijgen zou op de lange termijn heel nageslachtrijk kunnen zijn.) Het mag even zo lijken dat het beter is om aantallen kleinkinderen als relevante maat te nemen, maar steriele kleinkinderen vormen evolutionair gezien ook een doodlopende weg. Kortom de geschiedenis herhaalt zich. En fitness definiëren in termen van het aantal nakomelingen aan het einde der tijden is ook geen oplossing. Die is namelijk voor elk individu nul. Nu kijken populatiegenetici meestal niet naar de absolute fitness van individuen maar naar fitnessverschillen of fitnessverhoudingen. En dan is dit probleem redelijk oplosbaar (er blijven situaties denkbaar waarin deze oplossing ook niet werkt): kijk naar aantallen nakomelingen in de nde generatie, kies n zo klein mogelijk, maar als in generatie nummer n plus-een-beetje de fitnessverhouding plotseling drastisch verandert neem dan dan die generatie als basis voor je fimessbepaling. Maar dit alles terzijde. De uitwerking die ik hier gekozen heb is voor mijn doel in dit hoofdstuk goed genoeg.
bijlage 1 Lumsden en Wilson
Lumsden en Wilson[227] hebben een theorie ontwikkeld over genetische en culturele co-evolutie. Wat cultuur voor hen precies betekent is niet duidelijk, maar ruwweg komt het neer op het hele scala aan zaken en gedragingen waaruit een organisme kiezen kan. Een individu kiest steeds uit een reeks alternatieven die hem door zijn omgeving geboden wordt. Het gaat daarbij om een grote verscheidenheid aan zaken: de beschikbare varianten van een bepaald type kledingstuk; verschillende technieken om voedsel te verwerven; verschillende manieren waarop je een woord kunt uitspreken; verschillende manieren waarop je een soortgenoot kunt benaderen; verschillende uitvoeringen waarin een stuk gereedschap voorhanden is; de religieuze stromingen die in een samenleving voorkomen. Al die varianten en vormen heten in de terminologie van Lumsden en Wilson cultuurgenen (‘culturgens’).
Elk lid van een culturele diersoort leeft in een poel (engels ‘pool’) van cultuurgenen. De samenstelling van die poel is kenmerkend voor de betreffende cultuur. Wat een individu doet hangt af van de cultuurgenen waaruit het kiezen kan en de voorkeuren waarmee het is behept. En door keuzes te maken beïnvloedt het de samenstelling van de hem – en zijn soortgenoten – omringende cultuurgenenpoel. De manier waarop die keuzes tot stand komen vormt dan ook een belangrijk onderdeel van de basis van de theorie van Lumsden en Wilson. Ze gaan ervan uit dat in het genoom een verzameling biologische processen is vastgelegd die samen de ontwikkeling sturen van wat zij de geest (‘mind’)[228] noemen:
… the genes prescribe a set of biological processes, which we call epigenetic rules, that direct the assembly of the mind. This assembly is context dependent, with the epigenetic rules feeding on information derived from culture and physical environment. Such information is forged into cognitive schemata that are the raw materials of thought and decision. Emitted behavior is just one product of the dynamics of the mind, and culture is the translation of the epigenetic rules into mass patterns of mental activity and behavior. …[W]e take account of the free-ranging activities of the mind and of the diversity of cultures created by them. Genes are indeed linked to culture, but in a deep and subtle manner (Lumsden en Wilson 1981: 2).
In dit verhaal spelen epigenetische regels een dubbelrol. Aan de ene kant leggen ze vast hoe de geest zich ontwikkelt, en aan de andere kant beschrijven ze de manier waarop de geest, eenmaal volgroeid, informatie verwerft en verwerkt. Hoe die twee rollen aan elkaar zitten is onduidelijk. Ze worden door de auteurs niet expliciet onderscheiden. Wél maken ze onderscheid tussen twee fasen in het informatieverwerkingsproces van de geest die kunnen worden gekarakteriseerd met respectievelijk primaire en secundaire epigenetische regels:
Primary epigenetic rules are the more automatic processes that lead from sensory filtering to perception. … [An example is the way our visual cortex is] constructed so as to facilitate a perception of four basic colors.[229] The secondary epigenetic rules act on color and all other information displayed in the perceptual fields. They include the evaluation of perception through the processes of memory, emotional response, and decision making through which individuals are predisposed to use certain culturgens in preference to others (ibid.: 36).
Alle epigenetische regels samen bepalen de voorkeuren die een individu zal hebben bij een gegeven scala van cultuurgenen. In principe kun je dus, gegeven de epigenetische regels van een individu, voor elk cultuurgen uit een verzameling alternatieven aangeven met welke waarschijnlijkheid dat individu zich dat cultuurgen zal eigen maken. Voorkeuren kunnen worden weergegeven in een gebruiksvoorkeurscurve (‘usage bias curve’), of kortweg voorkeurscurve.
Met behulp van een eenvoudig gedachtenexperiment laten Lumsden en Wilson zien wat een voorkeurscurve precies is en wat je ermee opschiet bij het denken over evolutie en cultuur:
Suppose that a naive population or a naive individual of a given genotype within a species was confronted with a set of culturgens …, an assortment of food items, an array of carpenter’s tools, a variety of alternative marriage customs to be adopted or discarded, … If the development of individual members of the society is genetically constrained in such a way that the same culturgen is selected each time, the transmission is said to be pure genetic transmission [Voor alle duidelijkheid: verschillende individuen kunnen dan wel een verschíllende voorkeur hebben] (ibid.: 7-9).
Volgens de auteurs is zuiver genetische overdracht een logische mogelijkheid, maar ook niet meer dan dat:
… a highly social species might conceivably evolve that would possess an advanced language and culture all of which is learned, and yet be able to transmit only one set of behaviors and therefore one culture (ibid.: 10).
Het tegenovergestelde van zuiver genetische overdracht is zuiver culturele overdracht. Die doet zich voor wanneer er van geen enkele voorkeur voor een cultuurgen boven een ander sprake is.
Epigenetic rules have evolved so as to remove all forms of bias in individual development that could result from peripheral sensory screening, internuncial cellular organization, or innate predisposition in deeper processes of cognition (ibid: 10).
Welk cultuurgen een individu van zijn soortgenoten zal overnemen wordt dan volledig bepaald door welk cultuurgen hem toevallig het eerst wordt voorgehouden, dus door wat in de omgeving waarin dat individu opgroeit gebruikelijk is. Volgens Lumsden en Wilson geloven veel beoefenaren van de sociale wetenschappen dat deze vorm van overdracht kenmerkend is voor de mens.
Tussen deze beide uitersten ligt een derde mogelijkheid: een combinatie van genetische en culturele overdracht, kortweg gen-cultuuroverdracht. Daarvan is sprake wanneer minstens twee cultuurgenen een zekere, maar niet even grote, kans hebben om door een individu te worden overgenomen. In een eenvoudig schema (afb. 1) geven zij de drie vormen van overdracht weer, en wel in de vorm van de bijbehorende voorkeurscurven.
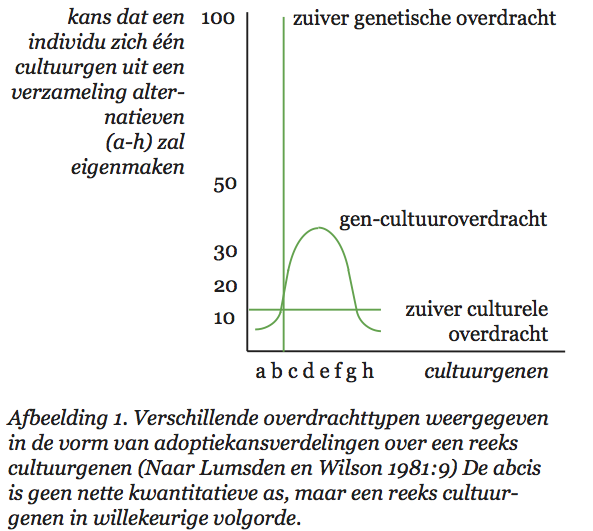
Volgens Lumsden en Wilson is gen-cultuuroverdracht voor mensen het meest waarschijnlijk. Zuiver genetische overdracht is niet meer dan een logische mogelijkheid, en zuiver culturele overdracht ligt om een aantal redenen evenmin voor de hand. In de eerste plaats zijn mensen niet voor alle stimuli even gevoelig, dus onze primaire epigenetische regels leiden al tot een zekere voorkeur. Bovendien is het evolutionair gezien ook te verwachten dat onze voorouders genetisch bepaalde[230] voorkeuren hadden:
Even if a species could somehow start with uniform epigenetic rules, the strategy would generally be unstable, leading in the course of gene-culture coevolution to the reappearance of nonuniform epigenetic rules. … Over a period of generations the population is unstable against invasion by genetic mutants that program epigenetic rules biasing individuals towards assimilation of relatively adaptive sets [of culturgens] (ibid.: 12-13).
Vervolgens beargumenteren Lumsden en Wilson dat mensen nog steeds onderworpen zijn aan genetisch-culturele co-evolutie. Wil dat zo zijn dan zal volgens hen aan vier voorwaarden moeten zijn voldaan.
Er zal sprake moeten zijn van voorkeursverdelingen van het gen-cultuur-overdrachtstype. Dat blijkt het geval.[231] Er zal sprake moeten zijn van genetische variatie in epigenetische regels. Dat daar sprake van is blijkt volgens Lumsden en Wilson uit het feit dat er voor zowat elke categorie van mentale activiteit, en van gedrag, die onderzocht is – kleuren zien, taalvaardigheid, en mate van extraversie, homoseksualiteit, kans op alcoholisme – genetische variatie is aangetoond. Verder zal er een verband moeten zijn tussen culturele gebruiken en voortplantingssucces. Ook dat blijkt het geval. Mensen vertonen allerlei gedragingen die gevolgen hebben voor hun levensverwachting en hun kans op voortplanting. Ten slotte zal er een moleculair en cellulair mechanisme moeten bestaan dat genen verbindt met onze cognitieve ontwikkeling. En hoewel we daarvan nog lang niet alle details kennen blijkt ook daaraan in principe voldaan. Hun theorie is, volgens Lumsden en Wilson, dus ook op de moderne mens volledig van toepassing.
In zekere zin is hiermee de basis van Lumsden en Wilsons theorie gegeven. Deze samenvatting is gebaseerd op de inleiding van hun eerste boek, Genes, Mind and Culture. De rest van dat boek besteden zij aan modelmatig onderzoek naar de consequenties van allerlei vooronderstellingen ten aanzien van de samenstelling van de cultuurgenenpoel van een bepaalde populatie, de erfelijkheid van epigenetische regels en het voortplantingssucces voortkomend uit allerlei combinaties van cultuurgenen. De wiskundige modellen die ze hiervoor ontwikkelen groeien me in ingewikkeldheid al na een paar pagina’s boven het hoofd. Ik kan de waarde daarvan dus niet schatten.[232] Mijn kritiek op hun visie berust dus eveneens uitsluitend op een analyse van de inleiding waarin ze de grondslagen voor hun theorie uiteenzetten.
kritiek op de uitgangspunten van Lumsden en Wilson
Een eerste aspect van het verhaal van Lumsden en Wilson dat bevreemding zou kunnen wekken is hun cultuurgen-idee. Dat suggereert dat ze cultuur opvatten als een verzameling discrete eenheden. Maar misschien is het goed om eraan te denken dat de genen die biologen soms als discrete eenheden opvoeren dat ook niet zijn, zonder dat daarmee nu meteen elk verhaal waarin ze zo wél optreden onzin wordt. Lumsden en Wilson zelf ontkennen met nadruk dat ze cultuur als een verzameling losse dingetjes hoeven op te vatten om over cultuurgenen te kunnen spreken:
… variation among relevant culturgens can be continuous … . The partitioning of such a continuum into categories is sometimes a practical step. We have taken care to show that the categories can be made as coarse or fine as required … and that continuous variation can be treated per se. We stress that culturgens are perceivable features of the integrated cultural system; their existence does not mean that culture is composed of discrete, isolated units (ibid.: 2).
Ik zei al eerder dat epigenetische regels een dubbelrol spelen in het verhaal. Ze beschrijven zowel de manier waarop genen de ontwikkeling van de geest sturen, als de manier waarop de geest selecteert uit de hem aangeboden cultuurgenenverzamelingen. In beide gevallen leggen de auteurs te veel nadruk op genetische en te weinig op omgevingsinvloeden. Laten we beide rollen een voor een bezien.
Volgens Lumsden en Wilson beschrijven epigenetische regels de invloed van genen op het zich ontwikkelende fenotype. Nu speelt de omgeving in die ontwikkeling eveneens een rol. En gezien de definitie van epigenese die Lumsden en Wilson geven onderkennen ze dat ook. Er zouden dus naast de epigenetische regels die de invloed van het genoom beschrijven ook epigenetische regels moeten zijn die de invloed van de omgeving op de ontwikkeling van het individu vastleggen. Maar die ontbreken in het verhaal. Een alternatief zou zijn dat epigenetische regels – als stuurders van de embryonale ontwikkeling – zelf omgevingsafhankelijk zijn, maar ook dat kwam kennelijk in de hoofden van de auteurs niet op. Op zich zou dit nog niet zo’n bezwaar zijn. De auteurs onderkennen (gezien hun definitie van epigenese) dat genen en omgeving beide een rol spelen in de embryonale ontwikkeling, en wie ze verwijt dat ze dat niet telkens opnieuw vermelden zoekt spijkers op laag water. Maar bij de tweede rol die epigenetische regels in hun verhaal spelen wordt het probleem nijpender.
Epigenetische regels, in een tweede betekenis, beschrijven de aangeboren voorkeuren van de geest. Kennelijk gaan Lumsden en Wilson ervan uit dat aangeborenheid iets met genetische bepaaldheid van doen heeft. Dat blijkt ondubbelzinnig uit de manier waarop ze hun overdrachtstypen (zie afb.1) presenteren. Ze onderscheiden drie mogelijkheden: het individu heeft honderd procent voorkeur voor een bepaald cultuurgen (genetische overdracht); het individu heeft in het geheel geen voorkeur (culturele overdracht); of het individu heeft een voorkeur die tussen nul en honderd procent in ligt (gen-cultuuroverdracht).
Dat lijkt een volledige opsomming van denkbare mogelijkheden: 100%, 0%, en alles daartussenin. Maar de schijn bedriegt. Er zijn meer variabelen in het spel dan de rigiditeit van de voorkeur. Het is bijvoorbeeld heel wel mogelijk dat een individu met een bepaald genoom, in welke omgeving het ook opgroeide, met een kans van 100% voor één bepaald cultuurgen zal kiezen, maar dat die voorkeur wel in verschillende omgevingen een ánder cultuurgen zou betreffen. De voorkeursverdeling in een bepaalde omgeving heeft dan altijd precies de vorm die volgens Lumsden en Wilson bij zuiver genetische overdracht hoort, maar er is duidelijk sprake van omgevingsvariatie en dus van omgevingsbepaaldheid in populatie-genetische zin. De mogelijkheid van een dergelijke omgevingsbepaalde ‘genetische’ overdracht (zie afb. 2.1) zien de auteurs over het hoofd. Op een zelfde manier zou er bij gen-cultuuroverdracht van omgevingsvariatie sprake kunnen zijn (afb. 2.2). Het is zelfs denkbaar dat de vórm van een voorkeurscurve van een individu afhangt van de omgeving waarin het zich ontwikkelde (afb. 2.3).
Kortom Lumsden en Wilson zien een zéé van mogelijkheden geheel over het hoofd! En dat komt uiteindelijk doordat ze geen rekening houden met de mogelijkheid dat voorkeuren en voorkeursverdelingen van individuen mede afhangen van de omgeving waarin die individuen leven.
Het door Lumsden en Wilson gecreëerde begrippenkader heeft vergaande consequenties. Het dwingt hen om bij ‘culturele overdracht’ meteen te denken aan, althans aanvankelijke, voorkeursloosheid. En dat wreekt zich vooral wanneer ze hun positie vergelijken met wat zij een kenmerkende stellingname van beoefenaars van de sociale wetenschappen achten.[233] Ze nemen aan dat die, wanneer zij de evolutionaire ontwikkeling van de mens bestuderen, vooral uitgaan van ‘culturele overdracht’: ‘…pure cultural transmission is what many social scientists have in mind when they interpret human evolution …’ (ibid.: 10). Dit soort overdracht houdt in dat ‘… all of the available culturgens are equally likely to be utilized. Epigenetic rules have evolved so as to remove all forms of bias in individual development …’ (ibid.: 10). Ze verwijzen naar onderzoek dat die visie ondersteunt. En ik neem ook graag aan dat iets dergelijks door sommigen wordt onderschreven.[234] Ik denk alleen dat Lumsden en Wilson dat idee net verkeerd interpreteren. De onderzoekers in kwestie geloven inderdaad dat genetische verschillen niet van doorslaggevend belang zijn voor een begrip van gedragsverschillen tussen mensen.[235] Ze gaan ervan uit dat de verschillen tussen pasgeboren mensen erg klein zijn of in elk geval dat die verschillen niet bepalend zijn voor cultuurverschillen. In die zin geloven zij in het belang van culturele overdracht. Maar daarmee is niet gezegd dat mensen geen aangeboren voorkeuren hebben (‘culturele overdracht’ in Lumsden en Wilsons zin des woords). Dat idee hangt niemand aan. Pasgeboren babies vinden zoet water lekkerder dan zout, verkiezen de harteklop van een mensenlijf boven de dreun van een drilboor en slapen rustiger in een warm en zacht bed dan op harde koude stenen. Niemand zal ontkennen dat hierbij sprake is van aangeboren ‘voorkeuren’. Maar uit het feit dat mensen aangeboren voorkeuren hebben volgt nog niet dat genetische verschillen relevant zijn voor een verklaring van cultuurverschillen en culturele veranderingen. We zouden heel goed een flink deel van onze aangeboren voorkeuren gemeenschappelijk kunnen hebben, en voor zover dat zo is zijn er helemaal geen verschillen tussen pasgeboren mensen die al dan niet genetisch bepaald kunnen zijn.
Kortom, in de theorie van Lumsden en Wilson spelen genen een veel te grote rol. Er is eigenlijk maar één plek waar aan het belang van de omgeving enige lippendienst wordt bewezen: in de definitie van epigenese. Daarnaast, of beter wellicht: ten gevolge daarvan, zien ze het fundamentele verschil tussen aangeboren kenmerken en genetisch bepaalde kenmerksverschillen over het hoofd. En dat op zich leidt er weer toe dat ze een vertekend beeld geven van de opvattingen van de sociale wetenschappers die ze als hun tegenstanders beschouwen. Ze maken daarmee een karikatuur van een redelijk alternatief voor hun eigen visie.
bijlage 2 Cavalli-Sforza en Feldman
Ook voor Cavalli-Sforza en Feldman[236] is cultuur een keuzekwestie. Na de geboorte wordt een individu geconfronteerd met soortgenoten, eerst en vooral de ouders, die zich op verschillende manieren gedragen. Uit dat scala van gedragsmogelijkheden kiest het – bewust of onbewust – een gedragspatroon dat het zich eigen maakt. Die keuze noemen de auteurs culturele selectie en het hele proces van bewustwording, keuze en verwerving, waarin leren een grote rol kan spelen, noemen zij culturele overdracht.
Cultural transmisions – the acquisition by one individual of a trait from another individual – may involve long and complex learning processes. … There are two stages in this transmission process, … The first stage is awareness, which requires the existence of a signal (via teaching and observation), and the second is acceptance (or learning). It is at the second stage that cultural selection may occur (Cavalli-Sforza en Feldman 1981: 62).
Na culturele selectie volgt een proces van natuurlijke (‘darwiniaanse’ ) selectie: ‘… Darwinian selection refers to the actual test by survival and fertility of the advantage of having or not having the trait’ (ibid.: 16). Cavalli-Sforza en Feldman brengen natuurlijke selectie in hun modellen in als een kans om tot voortplanting te komen – de fitness van een individu. Zulke kansen voor alle leden van een generatie leggen samen een deelverzameling van die generatie vast: de voortplantingsgemeenschap. Alle individuen die eenmaal door het natuurlijke-selectieproces zijn heengekomen krijgen vervolgens evenveel nakomelingen. Dat betekent dat een levenskrachtig individu dat is voorbestemd om weinig kinderen te krijgen in de modellen van Cavalli-Sforza en Feldman optreedt als een individu met een relatief lage kans om deel uit te maken van de voortplantingsgemeenschap. Het is even wennen, maar het werkt wel, omdat een zeer levensvatbaar individu met lage vruchtbaarheid uit populatiegenetisch oogpunt equivalent is met een individu dat een grote kans heeft voortijdig te overlijden maar dat eenmaal volwassen geworden wel veel nakomelingen krijgt.
Het opmerkelijke is nu dat in de modellen van Cavalli-Sforza en Feldman geen onderscheid wordt gemaakt tussen genetische en culturele overdacht. Daarin gaat het simpelweg over de fenotypische samenstelling van een populatie en de verandering daarvan in de loop van opeenvolgende generaties. Hun eenvoudigste model betreft een populatie waarin een bepaald kenmerk twee toestanden kent, en waarin de ouderfenotypen volledig bepalend zijn voor die van het nageslacht:
Let H and h represent the two states that the trait can take. There are then four possible mother-father pairs [mating types] … It is necessary to specify for each mating type the probability that the progeny be of type H or h. Denote by [the transition coefficients] b0, b1, b2, b3 probabilites (assumed constant) that an H child results from the matings h×h, h×H, H×h and H×H, respectively, where the mating specifies the mother × father pair with mother listed first. The frequencies with which these matings occur among adults at generation t in the population are p0 (t), p1(t), p2(t), p3(t), respectively, with
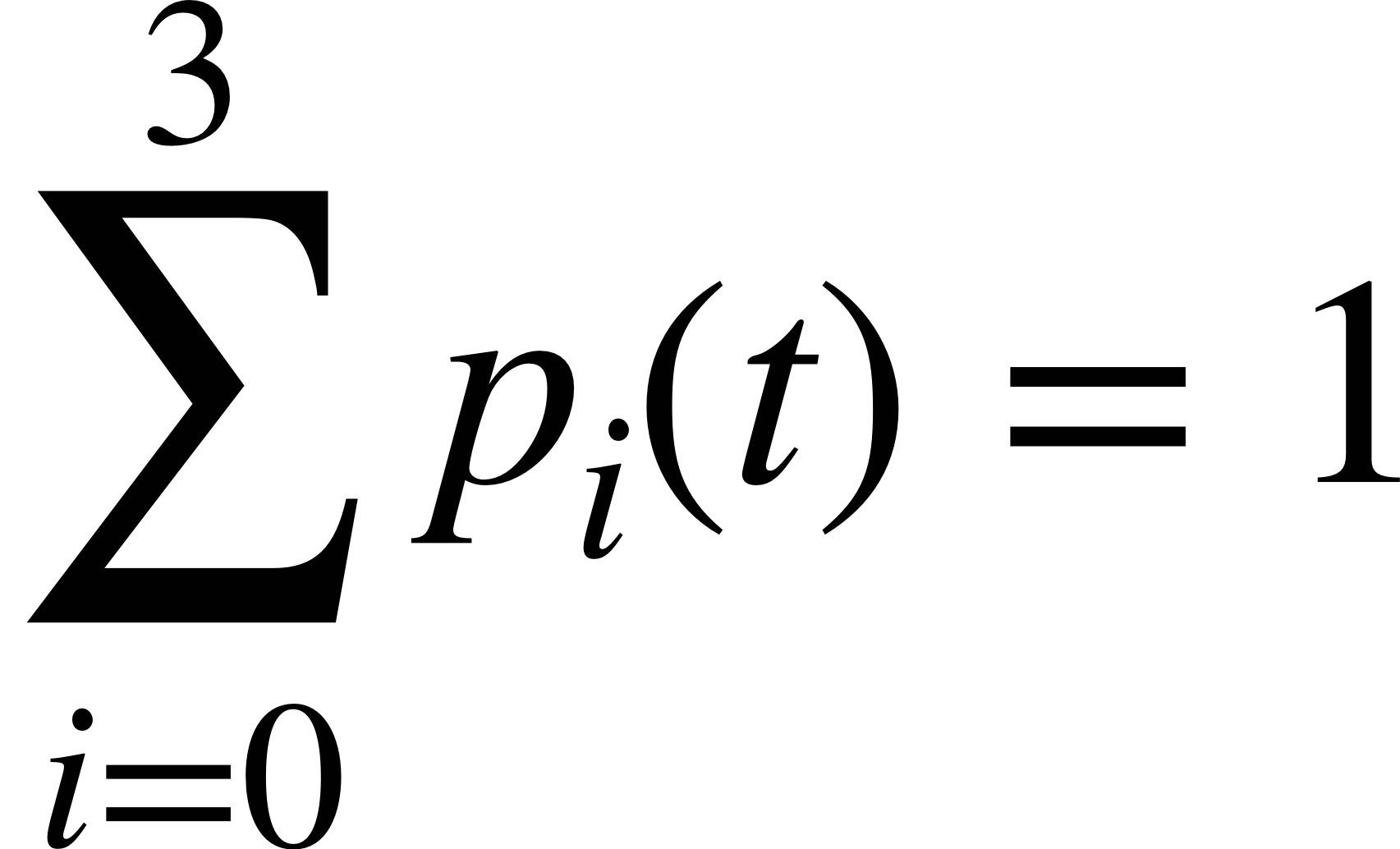
If the frequency of H in the adult individuals at generation t is ut, with
vt = 1-ut, that of h, then under random mating we have p0= vt2, p1 = utvt = p2, p3 = ut2, … The frequency of H in the offspring generation t+1 is
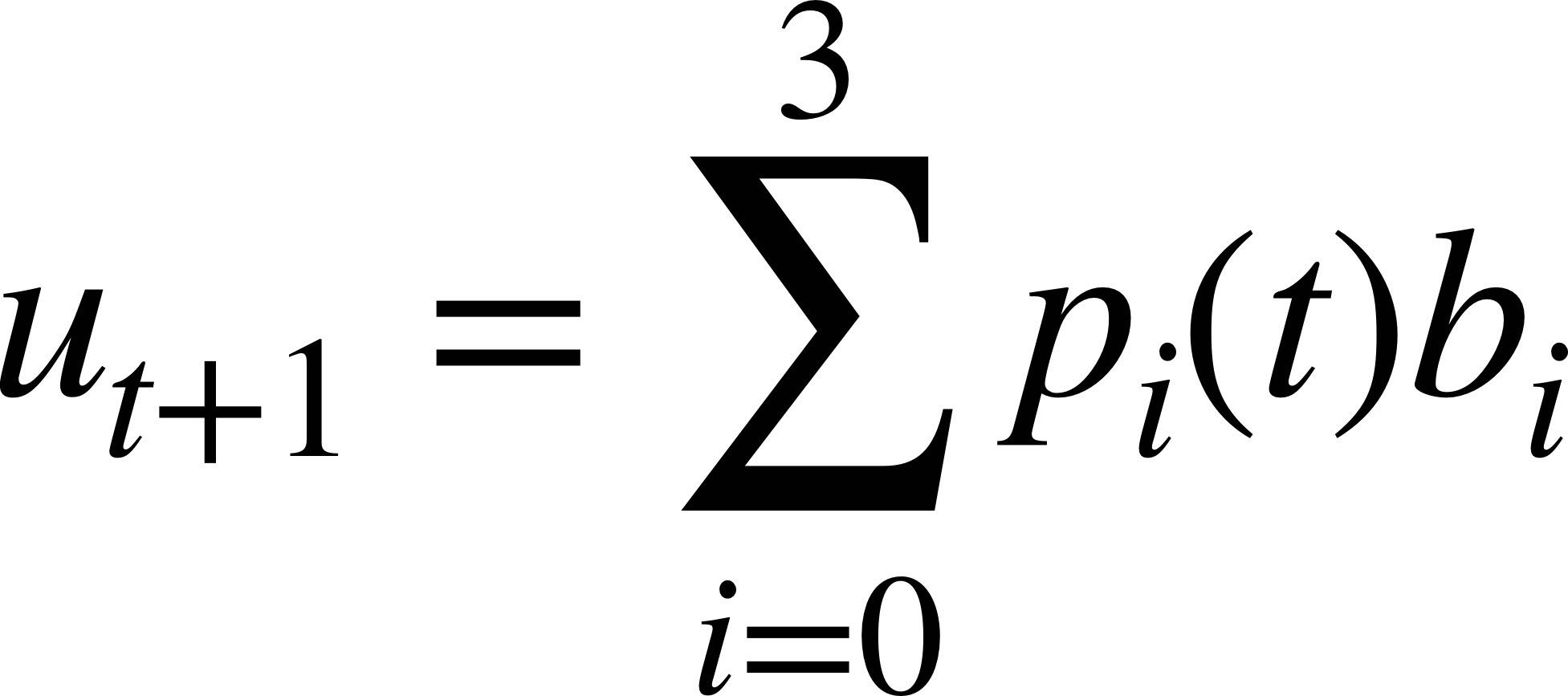
(ibid.: 78-79).
(Helaas sturen de kwadraatjes de regelafstanden enigszins in de war.)
In tabel 1 staan de belangrijkste elementen van het model op een rijtje.
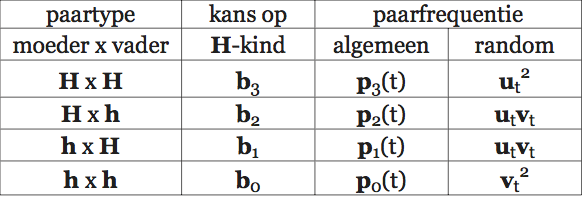
Tabel 1. De belangrijkste elementen uit het model voor culturele selectie waarin de fenotypen van het nageslacht volledig door die van de ouders worden bepaald (naar Cavalli-Sforza en Feldman 1981: 78, tabel 2.2.1).
Op basis van deze uitgangspunten ontwikkelen de auteurs modellen voor wat zij ‘culturele selectie’ noemen. Alleen waarom die selectie cultureel zou moeten heten is aan de uitgangspunten niet zo goed te zien. Ook situaties waarin het verschil tussen H en h volledig genetisch bepaald is, passen daar prima bij. De auteurs geven zelf een voorbeeld van een overdrachtscoëfficiëntencombinatie die zij aanduiden met ‘genetische overdracht’, zoals blijkt uit tabel 2.
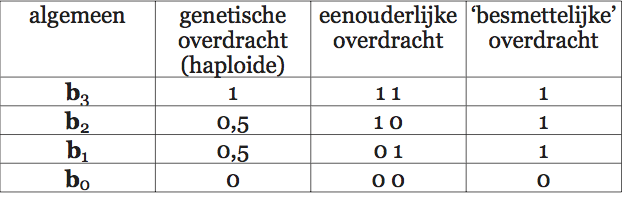
Tabel 2. voorbeelden van overdrachtscoëfficiënten van enkele denkbare overdrachtstypen (naar Cavalli-Sforza en Feldman 1981: 84, tabel 2.3.1).
Na dit eerste eenvoudige model volgen steeds ingewikkelder modellen waarin naast de ouders ook andere verwante en niet-verwante populatiegenoten als potentieel voorbeeld optreden. De auteurs onderzoeken de effecten van gerichte paarvorming, denk bijvoorbeeld aan situaties waarin H-individuen juist wel of juist niet voor elkaar vallen, van mutaties en allerlei vormen van natuurlijke selectie, en ze breiden hun modellen uit naar situaties waarin meer dan twee discrete toestanden van een kenmerk in het geding zijn.
Cavalli-Sforza menen dat hun modellen het eindelijk mogelijk maken het merkwaardige feit te verklaren dat culturele gebruiken soms leiden tot een verlaging van de levensvatbaarheid van de betrokken individuen. En ze geven een hele reeks voorbeelden van menselijke gedragingen die volgens hen zouden verdwijnen als natuurlijke selectie (in hun betekenis van het woord) de enige kracht zou zijn die tot frequentieverandering van fenotypen kon leiden. Mensen blijken regelmatig drugs te gebruiken, te roken, te veel en te vet te eten, te vasten, celibatair te leven, of hun lichaam allerlei verminkingen toe te brengen, denk aan besnijdenis van zowel mannen als vrouwen. En dat zijn allemaal zaken die hun voortplantingskansen nadelig beïnvloeden.[237] Een verklaring voor zo’n ongelukkig cultureel gebruik zou er volgens Cavalli-Sforza en Feldman als volgt uit kunnen zien (het voorbeeld is overigens van mijzelf).
Neem eens aan dat rokers gemiddeld minder kinderen krijgen dan niet-rokers. Als er geen sprake was van culturele selectie dan zou roken snel verdwijnen. Maar stel nu dat roken aanstekelijk werkt en wel zo dat alle kinderen waarvan minstens één ouder rookt zelf ook gaan roken (‘besmettelijke overdracht’, zie tabel 2). Er ontstaan dan drie soorten gezinnen: volledig abstinente ouderparen met een rijke en al even abstinente kinderschaar, gemengde huwelijken met minder kinderen die evenwel allemaal aan de nicotine zijn verslaafd, en vrijwel kinderloze gezinnen waarvan alle leden roken. Het verschil in kindertal tussen volledig abstinente ouderparen en ouderparen waarvan beide leden roken leidt op zich tot een afname van het aantal rokers, maar zolang uit gemengde huwelijken niet te weinig kinderen voortkomen kunnen die de afname van de rokersfrequentie volledig teniet doen. Rokers krijgen minder kinderen, maar die roken allemaal, terwijl de vruchtbaardere niet-rokers ten dele wel-rokend nageslacht krijgen. Op die manier zou culturele selectie de effecten van natuurlijke selectie kunnen weerstreven.
commentaar op Cavalli-Sforza en Feldman
In de modellen van Cavalli-Sforza en Feldman zijn fenotypische verschillen niet uitsluitend genetisch bepaald. Maar wat er zo typisch cultureel is aan wat zij toevoegen is onduidelijk. Het overdrachtsmechanisme uit mijn rokersvoorbeeld zouden zij ‘besmettelijke’ overdracht noemen. En die naam is heel aangemeten: elk kind waarvan zelfs maar een der ouders het betreffende kenmerk heeft krijgt dat kenmerk, en zo gaat dat vaak met een besmettelijke ziekte. Maar zo’n ziekte is dan ook meteen een eerste voorbeeld van een kenmerk dat met cultuur, in welke zin dan ook, niet zo veel te maken hoeft te hebben. Toch laat het voortbestaan van zulke ziekten zich met het model van Cavalli-Sforza en Feldman prima verklaren.
Wat is precies natuurlijke selectie in Cavalli-Sforza en Feldmans zin des woords? Kennelijk denken zij aan verschillen in nakomelingental tussen individuen met een verschillend fenotype. En dat noopt tot voorzichtigheid. Natuurlijke selectie wordt soms gedefinieerd in termen van verschillen in voortplantingssucces tussen individuen met een verschillend genotype. Zúlke verschillen zullen er meestal toe leiden dat het vruchtbaarste genotype in frequentie toeneemt, al zijn er uitzonderingen die we hier buiten beschouwing kunnen laten.[238] Alleen als een fenotypisch verschil direct samenhangt met een genotypisch verschil is er reden om te verwachten dat het vruchtbaarste fenotype in frequentie zal toenemen. Alleen als bijvoorbeeld het verschil tussen rokers en niet-rokers genetisch bepaald was, zou roken verdwijnen als het tot een relatief laag voortplantingssucces leidt en een universeel kenmerk worden in het tegenoverliggende geval. Het feit dat al die curieuze menselijke praktijken waarover Cavali-Sforza en Feldman zich verbazen niet verdwijnen is dus wellicht heel eenvoudige te verklaren, zelfs als we aannemen dat die praktijken inderdaad nadelig zijn voor het te verwachten kindertal.[239] Het zou kunnen gaan om kenmerken die niet vatbaar zijn voor natuurlijke selectie omdat er tussen rokers en niet-rokers, celibatiaren en niet-celibatairen, besnedenen en onbesnedenen, van genetisch bepaalde verschillen geen sprake is.
Er is geen enkele reden om bij de verklaring van praktijken die uit reproductief oogpunt nadelig lijken meteen aan ‘cultuur’ te denken. Zulke praktijken zijn waarschijnlijk helemaal niet zo zeldzaam. Bij alle organismen vinden we kenmerksverschillen die niet genetisch bepaald zijn. En in al die gevallen is het mogelijk dat de ene variant tot een groter reproductief succes leidt dan de andere. Cavalli-Sforza en Feldman hebben die mogelijkheid kennelijk over het hoofd gezien. Ook zij dragen cultuur aan om een probleem op te lossen dat ontstaat doordat het belang van omgevingsvariatie wordt onderschat.
Noten
[1] Wilson 1975. Zie de literatuurlijst voor meer gegevens.
[2] Er is bijvoorbeeld veel discussie over de vraag op welk niveau natuurlijke selectie plaats vindt. Gaat het bij selectie om een verschil in overlevingskans tussen genen, tussen organismen, tussen kleine of grote groepen van organismen? (Zie § 3.1.) Die vraag speelde al toen Wilson zijn Sociobiology schreef. Nergens blijkt echter dat hij zich met die vraag heeft beziggehouden. Wat hem betreft kan het blijkbaar allemaal, en wel net naar het de toevallige onderzoeker uitkomt.
[3] To sanctify a procedure or a statement is to certify it as beyond question and imply punishment for anyone who dares to contradict it. This extreme form of certification, the heart of all religions, is granted to the practices and dogmas that serve the most vital interests of the group. The individual is prepared by the sacred rituals for supreme effort and selfsacrifice . … Deus vult was the rallying cry of the First Crusade. God wills it, but the summed Darwinian fitness of the tribe was the ultimate if unrecognized beneficiary’ (Wilson 1975: 561). Wilsons verklaring is niet erg volledig. Hij vertelt wel wat zijns inziens het voordeel van godsdienst is, maar hij zegt er niet bij hoe precies godsdiensten zich ontwikkelden, en hij heeft het niet over groepen die wel en groepen die niet aan godsdienst deden.
[4] ‘Functie’, bijvoorbeeld, heeft in de biologie een heel specifieke betekenis. Het heeft betrekking op het selectief voordelige effect van een kenmerk en verwijst naar ontstaanswijzen. Wat een organisme nú doet met het onderdeel waarvan de functie in het geding is doet niet zo ter zake. Mijn vingers zijn bij uitstek geschikt om mee te typen, maar biologisch gezien is dat zeker niet hun functie. Een theoloog die stelt dat de functie van een of andere rituele handeling is dat die de aandacht op het Opperwezen gericht houdt hoeft dus niet per se ruzie te krijgen met een bioloog die beweert dat de functie van dat ritueel gelegen is in het creëren van een groepsidentiteit, zeker niet zolang de eerste bij functie denkt aan het nuttige effect van iets.
[5] Zie hoofdstuk 5 voor uitgebreid commentaar.
[6] Zie Huis in ’t Veld 1982.
[7] Zie Voorzanger en Sloep 1986.
[8] Zie de bijdrage van Van der Steen in Houtman e.a. 1986, voor een overzicht van de problemen die hier spelen.
[9] ‘Historisch’ betekent hier: betrekking hebbend op een in vele generaties gerekend verleden, en niet slechts – zoals de gewoonte onder historici dat wil – op het deel daarvan waaruit wij geschreven documenten bezitten.
[10] Uiteraard geldt dat voor willekeurig welke andere soort ook.
[11] Van Hooff wijst er terecht op dat dit zeker niet de mening van elke sociobioloog is. Met instemming verwijst hij naar Dawkins’ Selfish gene waarin deze ‘… stelt dat de mens het inzicht heeft verworven, dat het hem mogelijk maakt zijn eigen ontwikkeling te sturen, sociale contracten te ontwikkelen, ja zelfs zijn evolutie te richten – als hij dat wil’ (Van Hooff 1981: 35). Nu suggereert Van Hooff eerder (op dezelfde bladzij) dat de ‘vrije wil’ een begrip is dat alleen ‘… betekenis heeft in de persoonlijke ervaring.’ en dat … de ervaring een vrije keus te doen [niet inhoudt dat] het gedrag niet bepaald is en de keus niet verklaard kan worden!’ Ik waag mij niet aan een bespreking van de ‘vrije wil’, maar ik ontkom er niet aan mij af te vragen of het inzicht en de mogelijkheden die we volgens Dawkins en Van Hooff hebben geen inzicht en mogelijkheden zijn in een wel erg overdrachtelijke betekenis van deze woorden.
[12] Ik zal soms de termen ‘wetenschap’ gebruiken als ik het eigenlijk over ‘natuurwetenschap’ zou moeten hebben. De macht der gewoonte.
[13] Zie Van der Steen 1984a voor een helder overzicht van de problemen die hier spelen.
[14] Leaky en Lewin 1978 vermaken zich over verhalen van de !Kun San (een volk dat vroeger bosjesmannen werd genoemd) die wel weten wat allerlei dieren doen, die daar ‘zelfs’ (sic) een kennis van hebben die die van sommige westerse biologen evenaart, maar die, volgens de auteurs, volstrekt niet begrijpen waarom ze dat doen. Leeuwen zouden vooral jagen op een bepaalde gazellensoort omdat ze het vlees daarvan ‘lekker’ vinden. En ze begraven delen van hun buit om die te ‘verbergen’ voor hyena’s. Simplisten, die !Kun San! En zó onwetenschappelijk.
[15] Er bestaat een duidelijke parallel tussen de methodologie en de ethiek, en men zou in navolging van wat ethici doen onderscheid kunnen maken tussen een beschrijvende methodologie – wat doen wetenschappers als ze wetenschap bedrijven – en een normatieve methodologie – wat behoren ze te doen en waarom (Zie Sloep 1986). In dit boekje speelt de normatieve methodologie een hoofdrol.
[16] Voor een – op de biologie toegesneden – overzicht van methodologische criteria zie Van der Steen 1982, een beknopter overzicht met een duidelijke schets van onderlinge relaties geeft Sloep 1983: 5-13.
[17] Het woord ‘meerzinnig’ staat niet in Van Dale. Dubbelzinnig komt het dichtst bij wat ik hier bedoel, maar dat roept me net te veel associaties op met vette knipogen. Bovendien kan een meerzinnig begrip meer dan de door ‘dubbelzinnigheid’ gesuggereerde twee betekenissen hebben. Troost, tot slot, voor taalpuristen: als we ons allemaal uitsluitend beperken tot woorden die in woordenboeken staan kan onze woordenschat alleen maar langzaam kleiner worden.
[18] Zie Musschenga 1981 voor een uitgebreidere bespreking van dit punt.
[19] Formeel gezien gaat het bij meerplaatsigheid altijd om predicaten. Men spreekt in dit verband ook wel over ‘relationele begrippen’.
[20] Binnen de biologie zijn het vooral de evolutiebiologie (zie bijvoorbeeld: Ruse 1973; Hull 1974; Kitcher 1982; Sober 1984; Van der Steen 1982), meer in het bijzonder de sociobiologie (Ruse 1979; Kitcher 1985) waaraan wetenschapsfilosofen aandacht geven. Methodologische studies rond vakken als fysiologie, microbiologie, moleculaire genetica ontbreken vrijwel geheel. Dit kan de indruk wekken dat methodologische problemen zich uitsluitend voordoen in takken van biologie die vaak als ‘zacht’ worden ervaren. Het zou de moeite lonen om na te gaan in hoeverre die indruk terecht is.
[21] Bovendien zou onderzoek dat aan zo veel mogelijk methodologische eisen voldoet wel eens behoorlijk tijdrovend kunnen zijn, en dat past niet zo in ons tijdsgewricht.
[22] Zie bijvoorbeeld Van der Steen 1982 en 1984a.
[23] Om misverstanden te voorkomen: ik ben me ervan bewust dat er niet één algemeen geaccepteerde canon van methodologische criteria bestaat. Over zeer veel criteria bestaan forse meningsverschillen. Wat precies een theorie is, hoe we toetsbaarheid moeten opvatten, wat de bruikbaarheid van inductieve redeneringen is, is onderwerp van discussie, en zo is er vast nog veel meer. ‘Wetenschap’ staat dus voor een klasse van verzamelingen van methodologische criteria. Wat ik betoog is dat het wenselijk is die klasse zo klein mogelijk te houden. Hoe groter zij is, hoe minder het zegt wanneer een bepaalde uitspraak ‘wetenschappelijk’ heet. Het gaat me er dus niet om de verzameling toegestane methoden voor kennisverwerving in te perken, maar alleen de deelverzameling daarvan die we wetenschap noemen. Gewoon omdat het woord wetenschap daar preciezer en veelzeggender door wordt. (En ten overvloede: de veelzeggendheid waar het hier om gaat is geen kwestie van belang maar van duidelijkheid.)
[24] Voor een indruk van de eerste fase van dat debat in Amerika, de bakermat van de sociobiologie, leze men Caplan 1980; een eerste ronde van een debat in Nederland vond zijn neerslag in De Waal 1981.
[25] Zie Barker 1981.
[26] Alleen al vanwege die heftige en tendentieuze reacties lag mijn sympathie aanvankelijk bij de zwaar geplaagde sociobiologen. En hoewel ik hoe langer hoe minder heil zie in met name de Wilsoniaanse benadering, blijf ik van mening dat niets een rechtvaardiging kan vormen voor kritiek die gebruik maakt van tendentieuze inlegkunde en verdraaide citaten (zoals Allen et al 1978 in Amerika, en b.v. Huis in ’t Veld 1982 in Nederland, zie mijn reactie op Huis in ’t Velds verhaal, in Intermediair 18(30): 33, van 30 juli 1982).
[27] Darwin 1871.
[28] Zie Hull 1974, Ruse 1982 en Barker 1981 voor historische details.
[29] Bijvoorbeeld Morris 1967.
[30] Zie bijvoorbeeld zijn ideeën over verschillen in agressie tussen verschillende groepen mensen, in Lorenz 1966.
[31] ‘There is a close analogy between a human body invaded by a cancer and a nation inflicted with subpopulations whose inborn defects cause them to be social liabilities. Just as in cancer the best treatment is to eradicate the parasitic growth as quickly as possible, the eugenic defence against the dysgenic social effects of afflicted subpopulations is of necessity limited to equally drastic measures’ (Lorenz, in een vertaling van Chase, geciteerd in Barker 1981: 79).
[32] Barash 1979; Dawkins 1976.
[33] Boon en Smit 1985 gaan uitvoerig op dit verschil in, al is hun analyse niet helemaal bevredigend (zie voor commentaar: Voorzanger en Sloep 1986).
[34] Zie voor voorbeelden: Freeman 1980; Rose 1980; Simon 1980; Washburn 1980, Den Hartog 1981; Schuyt 1981.
[35] ‘… scientific discussions of human behavior manifest an uncanny resistance to moving beyond polarized debates about the role of nature vs. nurture, heredity vs. environment, or, to cite a more recent version of the same theme, biology vs. culture. The remarkable persistence of these categorizations bodes ill for those well-intentioned thinkers of various persuasions who hope to abandon these sterile labels once and for all’ (Caplan 1980: 100).
[36] Het gaat hier dus niet om een precieze weergave van wat auteurs feitelijk doen, maar om een schets van een analyseschema dat op de achtergrond meespeelt en dat de manier van denken bepaalt.
[37] Dit denken in parallel geschakelde tegenstellingen lijkt een diepgewortelde menselijke behoefte. In discussies over de aard van verschillende wetenschappelijke benaderingen wordt vaak onderscheid gemaakt tussen reductionistische en holistische, verklarende en begrijpende, machtgerichte en respecterende benaderingen. Dat zou acht verschillende benaderingswijzen moeten opleveren maar men doet vaak of er maar twee mogelijk zijn: de natuurwetenschapelijke die dan reductionistisch, verklarend, en machtgericht heet, en de sociaalwetenschappelijke die holistisch, begrijpend en respecterend zou zijn (zie Voorzanger l984a). In Boon en Smit 1985 gebeurt iets dergelijks met aan de ene kant: biologie, rationalisme, nature, aanleg, en aan de andere kant: sociale wetenschappen, empirisme, nurture, omgeving.
[38] En evenmin trouwens om dieren een aparte plaats te geven als het om menswetenschappelijke benaderingen gaat. In mijn betoog is dat geen hoofdpunt, maar ik vraag me wel eens af wat er gebeuren zou als sociologen of antropologen zich serieus met onderzoek aan dieren bezig gingen houden. Ik vermoed dat dat onze kijk op dieren drastisch zou kunnen veranderen.
[39] Bonner 1980 definieert cultuur in termen van aangeleerd gedrag.
[40] Lehrman 1970 en Dawkins 1982 kritiseren deze metaforen.
[41] Schellekens bijvoorbeeld spreekt over het uitwisselen van ‘eigenschappen’ in plaats van stukjes DNA als hij de mogelijkheden van recombinant-DNA-technieken bespreekt: ‘… het is nu mogelijk eigenschappen van een kangoeroe in een plant te zetten of eigenschappen van een tomaatplant in een olifant’ (Schellekens 1985: 11).
[42] Op zich is dat natuurlijk één stap in de goede richting. De verleiding om te denken dat er voor élk kenmerk één gen bestaat (en omgekeerd) ben je daarmee kwijt. En voor wie mocht denken dat biologen dat al lang niet meer denken, een citaat: ‘Jedem Merkmal kan man als Träger der Information ein Gen zuordnen’ (Schiegel 1969: 330).
Welnu dat kan natuurlijk, maar zinnig is het niet. Nu is Schlegel microbioloog, en microbiologische beschrijvingen van bacteriën bestaan vaak voor een groot deel uit een opsomming van enzymen die die bacteriën maken of kunnen maken. En bij elk enzym ‘hoort’ inderdaad een stukje DNA. Onbegrijpelijk is Schlegels stelling dan ook niet, maar bij kenmerken als de vorm van een bacterie, de structuur van z’n celwand, z’n delingssnelheid, past helemaal geen gen als informatiedrager. Weliswaar maakt Schlegel (op blz. 331) onderscheid tussen genotype en fenotype, maar hij doet het voorkomen alsof die zich verhouden als het organisme in aanleg – de blauwdruk dus – en het organisme zoals zich dat feitelijk ontwikkeld heeft.
[43] Ook Wilsons epigenetische regels horen in dit rijtje van misleidende metaforen thuis (zie bijlage 1).
[44] Volgens Faber 1986 kunnen we nog steeds spreken van het ‘dogma’ in de embryologie ‘… dat het DNA in de chromosomen een programma zou bevatten voor de embryonale ontwikkeling . … Het fenotype zou de min of meer directe ‘expressie’ zijn van het genotype’ (1986: 242). Faber betoogt vervolgens (blz. 243) dat het bouwplan voor het organisme niet in het DNA ligt maar geprogrammeerd is in de drie-dimensionale structuur van de eicel, met name die van het cytoplasma. Hij acht het DNA alleen van belang omdat het de moleculaire structuur van de bouwstenen van het zich ontwikkelende individu bepaalt. Alleen in die zin bevat het DNA informatie, maar het is het cytoplasma dat bepaalt hoe en wanneer die informatie wordt gebruikt. Als beschrijving van de machtsverdeling tussen celplasma en celkern zou dat wel eens heel reëel kunnen zijn. Met één element in Fabers verhaal ben ik intussen niet tevreden: volgens hem kán het DNA de informatie die benodigd is voor de embryogenese niet bevatten omdat het een ééndimensionale structuur is, terwijl embryogenese in zekere zin een vierdimensionaal proces is. Hij ziet niet hoe de ‘transpositie van één tot vier dimensies’ zou moeten verlopen, en stelt dat daar een logisch probleem ligt. Dat zie ik niet in. Wiskundigen hebben er geen moeite mee met eendimensionale formules driedimensionale lichamen te beschrijven. Logische barrières zijn er dus niet.
[45] Vroon 1981.
[46] Wilson 1978.
[47] Daarmee vooronderstel je dat het mogelijk is de mate van omgevingsbepaaldheid van de kenmerken van een soort te kwantificeren, en wel zo dat interspecifieke vergelijking mogelijk is, en dat zie ik zo een-twee-drie niet.
[48] De huizen en straten in die steden uiteraard niet.
[49] In hun monografie over het cultuurbegrip geven Kroeber en Kluckhohn 1952 ca. 170 verschillende, vooral aan sociaalwetenschappelijke literatuur ontleende, definities van cultuur.
[50] Lever 1966 geeft een hele reeks voorbeelden van organismen die gereedschap gebruiken en hun omgeving veranderen.
[51] Zie bijvoorbeeld: Sahlins 1977, en Schuyt 1981.
[52] Bonner 1980.
[53] Zie bijvoorbeeld Meulenbelt 1984 en Noske 1984. Noske, van wier verhaal ik de teneur overigens van harte onderschrijf, baseert een deel van haar redeneringen op de tegenstelling tussen biologisch en sociaal, en daarin kan ik haar niet volgen.
[54] Deze manier van denken werd voor het eerst duidelijk uiteengezet door August Comte, de vader van het positivisme (zie Edwards 1967 deel 2:174-175), en is bijvoorbeeld zeer ver uitgewerkt in de (reformatorische) Wijsbegeerte der Wetsidee met zijn vijftien zinkernen (zie Kalsbeek 1970: 27-29). Ik denk zeker niet dat alle denken over niveaus onzin is. Maar het probleem is dat er geen andere manier lijkt te zijn om niveaus van elkaar te onderscheiden dan door alle op een bepaald moment toevallig beoefende wetenschappen te nemen en die met enig kunst- en vliegwerk op een eendimensionale rij te zetten (zoals in de Wijsbegeerte der Wetsidee gebeurt). Voor iemand die niet gelooft in de mogelijkheid een vak tot een ‘onderliggend’ vak te reduceren, is dat wellicht een acceptabele procedure, maar een ontologische antireductionist (en dat zijn aanhangers van de WdW) zou hierdoor toch aan het denken moeten worden gezet.
[55] Voor voorbeelden van deze benadering zie Wilson 1978 en Barash 1979.
[56] Voor in de vorm van wiskundige modellen uitgewerkte theorieën over genetische en culturele co-evolutie zie Lumsden en Wilson 1981; Cavalli-Sforza en Feldman 1981; Boyd en Richerson 1985. Lumsden en Wilson presenteren hun uitgangspunten en werkwijze in Lumsden en Wilson 1982, en een overzicht van hun ideeën voor gewone mensen, dus zonder mathematische hokus-pokus, in Lumsden en Wilson 1983. Lopreato 1984 ontwikkelde een benadering die nauw met die van Lumsden en Wilson verwant is, maar zonder die in wiskundige modellen te gieten. Bonner 1980 geeft een schets van de evolutie van cultuur waarin het vermogen om te leren (al bij eencelligen aanwezig) en het vermogen om te onderwijzen noodzakelijke voorwaarden en zelfs in zekere zin voorstadia van cultuur zijn. Kern van zijn betoog is de geleidelijke ontwikkeling van culturele vermogens (evolutie van cultuur) en niet de ontwikkeling van b.v. menselijke culturen in de tijd (culturele evolutie). Dawkins 1976 en 1982, Maynard Smith 1982a, en Maxwell 1984 doen suggesties voor en geven aanzetten tot de ontwikkeling van theorieën over culturele evolutie.
[57] Een voorbeeld: ‘… we can understand the evolution of noncultural organisms as a process that transforms the distribution of genotypes from one generation to the next, because in a noncultural species each individual’s phenotype depends only on its genotype and the environment in which it matures.’ (Boyd en Richerson 1985: 6, cursivering van mij).
[58] ‘Fertilized eggs develop into organisms with different phenotypes. Laws of developmental genetics describe how a fertilized egg with a given genotype will produce different phenotypes, depending on the environment in which it develops. Here genotypic properties are mapped onto phenotypic ones by the laws T1. Given an array of organisms in a generation, let us say all at the juvenile stage, natural selection and other evolutionary forces may modify the composition of that generation as it matures to the adult stage. This transition is described by laws (T2) that map phenotypic descriptions onto other phenotypic descriptions. The adults then create gametes that then join to form fertilized eggs. Again selection may play a role, this time in producing differences in reproductive success. Phenotypic properties of parents may thereby affect the genetic composition of the gamete pool from which the next generation of zygotes will be constructed. Here, the requisite laws of transformation T3 move from the phenotypic to the genotypic level. The composition of gametes to form zygotes is described at the genetic level by a set of principles (T4) that includes the Hardy-Weinberg law, and we are off and running again with a new generation of fertilized eggs’ (Sober 1984: 37. Sober baseert zich hier vooral op Lewontin 1974: 12-13). Om het verhaal niet nodeloos ingewikkeld te maken heb ik bij mijn weergave in de hoofdtekst de verwijzing naar wetten en naar fenotypische en genotypische ruimtes weggelaten.
[59] Zie bijvoorbeeld Uyenoyama en Feldman 1980.
[60] De modellen van Cavalli-Sforza en Feldman, die ik verderop zal bespreken, zijn daar een voorbeeld van.
[61] Een verandering van de genetische samenstelling van een populatie ten gevolge van verschillen in voortplantingssucces die niet samenhangen met fitnessverschillen. Vooral in kleine populaties kan bijvoorbeeld het overlijden van enkele individuen ten gevolge van een ongeluk dat om het even wie had kunnen overkomen, de genetische samenstelling van de populatie drastisch veranderen.
[62] En dat geldt zelfs voor de, zeer ingewikkelde, modellen van frequentie-afhankelijke selectie en van coëvolutie van verschillendsoortige populaties (zie Falconer 1981 en Roughgarden 1979). Bij frequentie-afhankelijke selectiemodellen houdt men weliswaar rekening met het feit dat het voortplantingssucces van een individu kan afhangen van de samenstelling van de populatie, die op zich weer verandert onder invloed van selectie, en in zulke modellen is de omgeving van afzonderlijke individuen dus niet langer constant, maar dat komt alleen tot uiting in veranderende selectiedrukken. Voor coëvolutiemodellen geldt iets dergelijks. De mogelijkheid dat een omgevingsverandering op zich kan leiden tot fenotypische veranderingen, ook zonder dat er genetisch iets verandert, wordt ook dáár genegeerd. Ter voorkoming van misverstand: mijn betoog is niet gericht tegen populatiegenetica in engere zin, maar tegen een ruim opgevatte evolutiebiologie die populatiegenetica centraal stelt.
[63] Redeneringen als deze vindt u bij Lumsden en Wilson 1981, Cavalli-Sforza en Feldman 1981 en Boyd en Richerson 1985.
[64] Voorbeelden: Bonner definieert cultuur als ‘… the transfer of information by behavioral means, most particularly by the process of teaching and learning’ (Bonner 1980: 10).
Vergelijk ook de volgende citaten:
‘What gives man an extraordinary supremacy, what has permitted him to evolve at an unprecedented speed, is the transmission of learned experiences by means of an articulate and reflexive language’ (Dobzhansky en Boesiger 1983: 100).
‘Humankind has evolved by means of two entirely different interacting processes. One process, based on changes in the genes, is responsible for bodily structure and our capacity for acquiring knowledge and culture.
The second process, which determines how we use these capacities, depends on transfer of information from one individual to another’ (Stebbins 1982: 369-370).
Dawkins (1976, 1982) bedacht de ‘meme’ als cultureel analogon van het gen, ‘… a completely non-genetic kind of replicator, which flourishes only in the environment provided by complex, communicating brains’ (1982: 109). Veel meer dan een speelsigheidje om het algemene idee van een replicator te illustreren was dit niet, Dawkins werkte zijn ideeën niet verder uit en hij twijfelt aan de bruikbaarheid daarvan voor de bestudering van culturele ontwikkelingen (zie Dawkins 1982: 112).
[65] Boyd en Richerson 1985 doen dat alleraardigst.
[66] ‘… variants acquired by individual learning and other common forms of phenotypic flexibility are lost with the death of the individual. … In contrast, culturally acquired variations are transmitted from generation to generation and, like genes, they are evolving properties of the population’ (Boyd en Richerson 1985: 4).
[67] Gupta en Lewontin 1982.
[68] Zie Trivers 1985 voor voorbeelden.
[69] Zie bijvoorbeeld Fossey 1984.
[70] Zie bijvoorbeeld de bijdragen van Campbell en anderen in Stent 1978.
[71] Sommigen zullen dat een onzinnige vraag vinden. Zij weten al dat het antwoord ‘niets!’ zal zijn. Ik ben het daar van harte mee eens, maar ik denk dat het niet alleen belangrijk is om gelijk te hebben, het is ook belangrijk om goede gronden voor dat gelijk aan te voeren.
[72] Dit klinkt misschien hoogdravender dan ik het bedoel. Ik ga er even van uit dat alle dingen die we doen, vanuit een ethisch oogpunt, kunnen worden opgedeeld in goede en slechte, egoïstische en altruïstische, rechtvaardige en onrechtvaardige. In evolutionair perspectief is geen van die indelingen relevant. Andere indelingen zijn dat wel: gedrag dat wel en gedrag dat niet zal worden weggeselecteerd, bijvoorbeeld. Maar dat is in ethisch opzicht weer niet zo’n relevant onderscheid.
[73] Hamilton 1964.
[74] Dat is dus precies de beperking waarvan ik in het vorige hoofdstuk betoogde dat het voor evolutiebiologen tijd werd om die los te laten.
[75] Dit geldt alleen voor niet-geslachtsgebonden genen van diploïde organismen, waarbij homozygotie zeldzaam is, en die leven in een niet te kleine populatie waarbinnen partnerkeuze ‘random’ is met betrekking tot genetische overeenkomst. En zo zijn er vast nog wel wat beperkingen.
[76] Omdat er geen Nederlands equivalent voor dit woord bestaat en de Engelse term in de biologie is ingeburgerd en een vrij precieze betekenis heeft zal ik die hier verder ook gebruiken.
[77] Soms doet men het voorkomen alsof verwantenselectie en ‘gewone’ selectie verschillende processen zijn die elk een rol spelen, maar als Hamilton gelijk heeft dan is gewone selectie, als men daarbij het simpele beeld op het oog heeft dat Hamilton onderuit haalde, geen alternatief, vanwege de denkfout die daarachter steekt. Er bestaat overigens een zee van verwarring rond het verwantenselectiedenken. Zie voor een eerste overzicht Dawkins 1979.
[78] Zie Wilson 1975: 448-455.
[79] Doordat mannetjes haploied zijn zijn de dochters van één vader wat het vaderlijke deel van hun genoom betreft identiek. Zie Wilson 1975: 416.
Vooral over dit en verwante voorbeelden hangt een sluier van verwarring. Zo doet men het soms voorkomen alsof het geweldige aantal kinderen van een bijenkoningin een compensatie vormt voor de onvruchtbaarheid van werkbijen, maar de meeste van die nakomelingen worden werksters, ze zijn dus steriel. Het moge duidelijk zijn dat een overmaat van doodlopende wegen niet echt een evolutionaire uitweg biedt. Wat telt is het succes van een koningin gemeten in aantal dochterkoninginnen en darren.
[80] Falconer 1981.
[81] Uyenoyama en Feldman 1980.
[82] Zie Wynne-Edwards 1963, en verwijzingen daarin.
[83] Zie voor voorbeelden Boorman en Levitt 1980.
[84] ‘As seen by present day evolutionists, Darwin’s scheme embodies three different principles …: 1. Different individuals in a population have different morphologies, physiologies and behaviors (phenotypic variation). 2. Different phenotypes have different rates of survival and reproduction in different environments (differential fitness). 3. There is a correlation between parents and offspring in the contribution of each to future generations (fitness is heritable). These three principles embody the principle of evolution by natural selection.’ (Lewontin 1970: 1).
[85] Lewontin schrijft wel vaker dat er sprake moet zijn van wat hij noemt ‘erfelijke variatie in fitness’. Ik vind dat enigszins raadselachtig. Het ligt zo voor de hand om te eisen dat de fenotypische variatie erfelijk moet zijn. Of zou Lewontin die nadruk op fitness leggen om te voorkomen dat men bij elke fenotypische variatie meteen aan selectie gaat denken?
[86] Ook populaties en soorten ‘stammen’ per slot van rekening ‘af’ van andere populaties en soorten.
[87] Ook bijvoorbeeld Wilson (1975: 106-107) verdedigt die visie, al treden bij hem opeens verwantengroepen op als bijzondere categorie op het groepsniveau. Dat laatste leidt tot debatten over de vraag of verwantenselectie inderdaad een vorm van groepselectie is. Die vraag laat ik hier graag liggen. Zolang niet duidelijk is wat je precies bedoelt met selectie heeft zo’n vraag weinig zin. Zie verder Van der Steen en Voorzanger 1984a. (De lezer make uit de overeenstemming tussen Wilson en Lewontin overigens niet op dat zij het verder over veel eens zijn. Lewontin is een gestaalde tegenstander van sociobiologische theorieën over de mens.)
[88] Dawkins 1976 en 1982. Zie vooral Dawkins 1982: 82-83 voor een weergave van zijn visie op selectie. Zie Van der Steen en Voorzanger 1984a voor uitgebreider commentaar op Dawkins’ benadering.
[89] Hoewel b.v. Hulls ideeën wel wat op die van Dawkins lijken (zie Hull 1980).
[90] De lezer make hier niet de fout te denken ‘ja, dat herken ik, al die stemmetjes in mijn hoofd die mij elk een andere kant op commanderen’. Ik bezondig me hier aan een manier van spreken die biologen soms erg eigen is: het vertalen van een ingewikkeld en onpersoonlijk verhaal in termen van beweegredenen van individuen. Dat is beeldspraak die men ook als zodanig dient te herkennen.
[91] ‘Hetzelfde locus’ zou hier de geëigende formulering zijn. Zie tweede voetnoot van 3.2.2.1.1 voor terminologische uitleg.
[92] Of preciezer: als het verschil tussen de wereld zoals die is en de wereld zoals die zijn zou als er op de plek van dat gen een ander gen zat.
[93] Zij het dan een theorie met veel haken en ogen. Zie mijn bespreking van Cavalli-Sforza en Feldman in hoofdstuk 2 en bijlage 2.
[94] Voor alle duidelijkheid: of een kenmerk in een populatie voorkomt hangt er natuurlijk niet alleen van af of het al dan niet wordt weggeselecteerd, maar bijvoorbeeld ook van de snelheid waarmee nieuwe dragers van dat kenmerk door genmutatie ontstaan.
[95] Er zijn ethologen die intenties opvatten als interne representaties van een nagestreefde toestand (Van Hooff, persoonlijke mededeling). Intentioneel gedrag is dan gedrag dat erop gericht is om de wereld zodanig te beïnvloeden dat er een toestand ontstaat die overeenkomt met die interne voorstelling. Een organisme dat daarmee bezig is zal steeds opnieuw nagaan in hoeverre de overeenkomst is gerealiseerd, en zijn gedrag aanpassen aan de aard en de richting van het waargenomen verschil. Zo’n benadering roept een aantal vragen op. In hoeverre draagt het postuleren van een ‘bedoeling’ bij aan ons begrip van het gedrag van een organisme? Creëer je daarmee geen inhoudsloze categorieën (zoiets als het ‘instinct’, waar de meeste ethologen om die reden niet meer over spreken)? In hoeverre sluit deze interpretatie van ‘intentie’ aan bij onze alledaagse interpretatie, of bij die waar bijvoorbeeld sommige ethici vanuit gaan? Deze en soortgelijke vragen zijn onderwerp van een uitgebreid en voorlopig onbeslist debat waarbij zeer veel verschillende wetenschappen in het geding zijn (zie bijvoorbeeld Toates 1986, Churchland 1986, Van der Steen en Thung 1988).
[96] Zie o.a. Hamilton 1964, Trivers 1971, Wilson 1975.
[97] ‘Spite’, zie Wilson 1975: 117. Volgens Wilson (1975: 119) zal kwaadaardigheid zeldzaam zijn omdat het ‘the intelligence to plot intrigue …’ vooronderstelt. Waarschijnlijk om dezelfde reden verwacht Barash (1979: 32) dat het bij dieren in het geheel niet voorkomt.
[98] ‘Cooperation’, zie, bijvoorbeeld, Kurland 1977.
[99] De meesten nemen niet eens de moeite om aandacht te geven aan mogelijke verschillen. Bertram 1982 constateert expliciet dat die verschillen zeer klein zijn, hij acht het ook ‘… desirable for the concept of altruism in biology to be similar to that used for human beings. … the closer the definitions the better, since one would hope that less confusion should then arise’ (blz. 252). (De tegenstelling tussen altruïsme in de biologie en altruïsme zoals dat voor mensen wordt gebruikt, vormt het zoveelste voorbeeld van het dichotome denken waar hoofdstuk 2 over ging.)
[100] Zie Musschenga 1981.
[101] Als we ‘altruïsme’ definiëren in termen van de gemiddelde effecten van een bepaald soort gedrag, en niet in termen van de feitelijke effecten van een zeer bepaalde handeling dan kunnen we het soort ongelukjes dat ik in mijn voorbeeld gebruik, negeren. Ik zou hier dan een voorbeeld moeten geven van gedrag dat goed bedoeld is maar dat doorgaans vooral negatieve effecten heeft. En dat zou misschien een erg vergezocht voorbeeld worden. Hoewel, welke beroepsgroep blijft gevrijwaard van beunhazen die met de beste bedoelingen steeds opnieuw meer versjteren dan ze goedmaken? Of zouden we die beunhazen zélf als te negeren ongelukjes moeten beschouwen?
[102] In navolging van Barnett 1980: 144.
[103] Verhalen over het onderscheid tussen het gedrag van katten en dat van honden, of over Jan die zich zo geheel anders gedraagt dan zijn broer Sander, gaan over gedrag in de eerste zin des woords. Wie vraagt wat Jan deed toen hij vanochtend te laat op school kwam, vraagt naar gedrag in de tweede betekenis. Voor die tweede betekenis bestaat in het Nederlands eigenlijk geen apart woord. Weliswaar kan het onderscheid dat ik op het oog heb goed worden weergegeven met het onderscheid tussen ‘het handelen’ en ‘de handeling’, maar als ik die begrippen hier zou gebruiken zou ik het verschil tussen handelingen en gedragingen dat in sociaalwetenschappelijke theorie wordt gemaakt, negeren: handelingen zijn bewuste gedragingen. Om misverstanden te voorkomen gebruik ik hier de termen ‘gedrag’ voor de manier van doen in het algemeen en ‘gedraging’ voor specifieke (reeksen van) uitingen van een individu. Zie verder 3.2.2.1.2 over ‘strategieën’.
[104] Zie Uyenoyama en Feldman 1980.
[105] Cactusmussen zijn vogeltjes die dankzij hun vaalgroene licht gepunte veren sprekend lijken op de jonge loten van de cactussen waartussen ze leven. Ze zijn dus, op zo’n cactus gezeten, prachtig gecamoufleerd, vooral wanneer ze hun veren rechtop zetten. Ten overvloede: cactusmussen bestaan niet.
[106] Genen stelt men zich voor als DNA-moleculen die in lange ketens aaneengeregen zijn. Die lange ketens zijn meestal afzonderlijk verpakt in een eiwithulsje en zo’n geheel van genenketen plus huIsje heet een chromosoom. Bij de meeste ‘hogere’ organismen komen chromosomen voor in paren. Overeenkomstige plekken op de beide leden van een chromosomenpaar noemt men een ‘locus’.
[107] Er is hier dus sprake van wat met een technische term ‘heterosis’ heet: de heterozygoot heeft de grootste fitness.
[108] Lever (persoonlijke mededeling) merkte hierbij terecht op dat dat in zekere zin nog niet precies genoeg is. Ten gevolge van mutaties tijdens het leven van een individu kan (een deel van) het fenotype anders worden dan het zonder die mutaties geweest zou zijn. Die mutaties hoeven niet ook de voortplantingscellen te bereiken, dat hangt er maar net van af waar en wanneer ze optreden. Je zult dus onderscheid moeten maken tussen het genenbestand dat het fenotype (en dus ook de voortplanting) beïnvloedt en het genenbestand dat bij reproductie kan worden doorgegeven (resp. het somatische en het kiembaangenoom).
[109] Terzijde: of ‘groene ogen’ één kenmerk is hangt af van het soort variatie in de populatie. Wanneer de kleur van de bovenste helft van de iris onafhankelijk van die van de onderste helft zou variëren, zou ‘groene ogen’, ondanks de talige suggestie van het tegendeel, twee kenmerken zijn. Een kenmerk is in zekere zin elk onderdeel van een organisme waarin het van andere organismen kan verschillen.
[110] Dominey 1984 geeft een heldere analyse van het strategiebegrip. Mijn omschrijving is aan hem ontleend.
[111] Aangenomen ten minste dat het mogelijk is om het gedrag van organismen in termen van een strategie te beschrijven. Je kunt zo’n begrip wel definiëren, maar daaruit volgt niet dat het gedefinieerde ook werkelijk bestaat. Het probleem is dat gedrag niet alleen afhangt van de omstandigheden nú. Soms doe ik bij een graad of tien Celcius een jas aan, soms doe ik bij die temperatuur mijn jasje juist uit, dat hangt ervan af hoe warm of koud het voordien was en in hoeverre ik me daarop gekleed had. Dat is nog wel te verwerken door niet naar omstandigheden sec maar naar omstandighedenveranderingen te kijken. Maar lastiger wordt het wanneer mijn reactie op een verandering van omstandigheden nu, op zich weer afhangt van wat me langer geleden overkomen is. Daarop voortbordurend zie je geleidelijk aan het risico opdoemen dat een nette strategiebeschrijving gaat bestaan uit een volledige opsomming van alle denkbare levensgeschiedenissen, die elk op zich gedetailleerd beschreven moeten zijn. En dat zou geen werkbaar materiaal zijn.
Voor één ding moeten we oppassen: een strategie is geen onderdeel van een organisme op de manier waarop een arm of een zwezerik dat is. Hij ‘zit’ niet ergens in het beest, in de hersenen of zo. Hij bestaat alleen op papier. Natuurlijk is dat wat een strategie beschrijft het gevolg van hoe een beest in elkaar zit, maar dan wel van het héle beest.
[112] Zie Maynard Smith 1982a.
[113] Door aan alle denkbare wederwaardigheden (verloren en gewonnen gevechten, tijd besteed aan weglopen, enzovoort) consequenties te verbinden in de vorm van winst- of verliespunten, en een aantal vooronderstellingen te doen ten aanzien van aantallen ‘duiven’ en ‘haviken’ in een populatie, ontmoetingskansen, winstkansen bij een ontmoeting van twee ‘haviken’, enzovoort, kun je berekenen welke strategie het meest profijtelijk is. Het aardige van dit vrij simpele model is het onverwachte resultaat dat ‘duiven’, als er niet te veel van zijn en als de consequenties van een verloren gevecht ernstig genoeg zijn, soms beter af zijn. Ten overvloede: het gaat hier natuurlijk niet om echte duiven en echte haviken, echte haviken zijn helemaal niet zo strijdlustig, zeker niet tegenover hun soortgenoten, en echte duiven kunnen soms behoorlijk tekeergaan, dáárvan ben ik regelmatig getuige.
[114] Zie Dawkins 1982: 179-194 voor een overzicht van gangbare interpretaties het fitnessbegrip.
[115] Beatty 1984: 191.
[116] Beatty 1984: 192.
[117] ‘… The number of offspring that it is physically disposed to contribute … in a particularly specified environment’ (Beatty 1984: 192).
[118] De zgn. ‘propensity’-interpretatie van ‘fitness’, zie Beatty 1984 en Sober 1984.
[119] In zekere zin verhouden die termen zich als het klimaat en het weer in ons dagelijks leven: het klimaat is relatief constant in de tijd en over een vrij groot geografisch gebied, terwijl het weer van minuut tot minuut kan veranderen en hier beduidend anders kan zijn dan een paar kilometer verderop. Het klimaat is een aspect van de omgeving waarin we leven, het weer van de omstandigheden.
Omgeving en omstandigheden zul je uiteindelijk moeten definiëren met betrekking tot een bepaald type organisme en een bepaalde vraagstelling. ‘In het bos’ is een aardig antwoord op de vraag in wat voor omgeving eekhoorntjes wonen, maar als de vraag betrekking heeft op boktorren die heel specifiek aan bepaalde boomsoorten gebonden zijn is het een veel te globaal antwoord, terwijl het voor reigers die zich in de stad ook prima staande kunnen houden een veel te beperkte aanduiding is. Voor een reiger is het bos meer iets dat je kunt rekenen onder de omstandigheden waaronder hij zich toevallig kan bevinden.
[120] Stel het kindertal van de i-de levensweg is ni de kans om die levensweg te volgen is pi, dan is de darwiniaanse fitness Σni.pi.
[121] Vergelijk Sobers definitie van Darwiniaanse fitness: ‘the mathematical expectation of the organism’s number of offspring’ (Sober 1984: 45). De manier waarop ik het probabilistische karakter van het fitnessbegrip hier heb uitgewerkt is vast niet de enige mogelijkheid. Ik heb gestreefd naar een uitwerking die technisch gedoe zo veel mogelijk vermijdt en die de lezer niet nodeloos doet verdwalen in een fancy formulepark. ( Zie opmerking direct na de literatuurlijst voor meer over het fitness-begrip).
[122] Ik denk soms dat je zou kunnen verdedigen dat de inclusieve fitness – mits gedefinieerd in termen van feitelijke kindertallen van het individu en al zijn verwanten – een heel aardige schatting van de darwiniaanse fitness van een individu zou kunnen opleveren. Maar dat zou verder doordacht moeten worden.
[123] Of, maar dat is een wel erg theoretische mogelijkheid, wanneer het genoom van het individu verandert. Voor de volledigheid: ik ga er dus van uit dat het fenoom van een individu alleen kan veranderen door een verandering van de omgeving en/of het genoom.
[124] Hoewel je met die conclusie, zoals Lever mij terecht zei, moet oppassen: er zijn samenlevingen waarin martiale lidtekens de kansen op de huwelijksmarkt kunnen vergroten, en veel verwondingen gaan gepaard met een stukje levenservaring dat op zich de levenskansen kan vergroten!
[125] Gedrag (1) van individu (2) is altruistisch tegenover individu (3). Zie verder 1.2.2, en Voorzanger 1983 en 1984b.
[126] Als je bij welstand niet al te benepen alleen aan materieel bezit denkt maar ook bijvoorbeeld aan informatie, dan is zoiets als waarschuwen voor een naderend roofdier ook altruïstisch in, in elk geval, deze zin.
[127] Volgens Uyenoyama en Feldman zou bioaltruïsme zelfs in termen van genotypen gedefinieerd móeten worden ‘… because fitness is a statistical property of the genotype rather than of the individual, such a definition [d.w.z. in termen van individuen] is necessarily incomplete’ (Uyenoyama en Feldman 1980: 381). Of ze gelijk hebben hangt af van het fitnessbegrip dat je hanteert. Gegeven de definitie van fitness die ik eerder gaf is spreken over de fitness van een individu geen enkel probleem. Maar ze zouden in die zin gelijk kunnen hebben dat de fitness van een genotype in evolutietheoretisch opzicht interessanter is dan die van een individu.
[128] Het probleem is hier dat je, denk ik, graag zou willen spreken over het gedrag dat de oorzaak is van kindertal(verm)-veranderingen, maar als gedrag een al dan niet directe reactie is op, gevolg is van, een ander gedrag is dat andere gedrag een net iets uiteindelijkere oorzaak van de kindertal(verm)-veranderingen in kwestie. Hier dreigt een oneindige regressie die alleen met gezond verstand te stoppen valt. Maar het gezonde verstand van de ene onderzoeker is dat van de andere onderzoeker niet.
[129] Voor de lezer die al paragrafen lang tevergeefs zit te wachten tot ik het nu eindelijk eens over Trivers’ ‘wederkerig altruïsme’ (Trivers 1971) ga hebben: het feit dat altruïsme in de tweede en de derde betekenis eigenlijk niets met elkaar te maken hebben is in zekere zin precies wat Trivers laat zien. Wederkerig altruïsme treedt op wanneer een individu iets voor een ander doet dat hem op de korte termijn schaadt maar dat later door die ander wordt goedgemaakt. Gegeven mijn eerste twee definities is dat dus gewoon altruïstisch gedrag, gegeven mijn derde definitie is het dat gewoon niet. ‘Wederkerig altruisme’ is dus een wat verwarrend begrip. Maar dat neemt niet weg dat de mogelijkheid dat een organisme invloed heeft op wat hij op de lange termijn van anderen verwachten kan natuurlijk heel interessant is.
[130] Net zo goed als individuen met S2 altruïstisch zouden zijn vergeleken met individuen voor wie de kansverdeling bijvoorbeeld 90% - lO% zou zijn!
[131] En wanneer dat niet gebeurt is er alle reden om op zoek te gaan naar de oorzaken daarvan. Die zouden bijvoorbeeld gelegen kunnen zijn in de genetische structuur (denk aan heterosis) of de frequentieafhankelijkheid van fitnessen. Wat dat betreft geldt hier hetzelfde als bij elk ander kenmerk.
[132] Tenzij je aanneemt dat sommige of alle gedragingen die altruïstisch zijn in de zin van de eerste twee definities voortkomen uit strategieën die met een simpele genetische verandering omgezet zouden kunnen worden in strategieën die tot vermijding van het betreffende gedrag zouden hebben geleid en die een hogere fitness zouden hebben, en tevens dat elke denkbare genetische variant ook werkelijk ontstaat. Die eerste veronderstelling is gegeven onze huidige kennis van de relatie tussen genen en gedrag op geen enkele manier te toetsen en de tweede berust op het simpele geloof dat de natuur alles wat logisch gesproken tot de mogelijkheden behoort ook daadwerkelijk en onmiddellijk uitprobeert. En wat ik daar van denken moet weet ik werkelijk niet.
[133] De Graaf 1972.
[134] Soms gaat het daarbij om moraaltheologen.
[135] Ik denk dat het niet kan, maar die overtuiging komt voort uit maar één overweging: ik zou werkelijk niet weten hoe je uit feiten normen kunt afleiden op een manier die overtuigend is voor mensen die die normen niet al aanhangen. Pogingen als die van Searl, Gewirth, Richards (zie verderop), om te laten zien dat het wel kan, verbazen me voornamelijk.
[136] Zie Mackie 1977; Hudson 1983; Hubbeling en Veldhuis 1985.
[137] Je zou dit moreel objectivisme kunnen noemen. Volgens Mackie is een objectivist noodzakelijkerwijze ook altijd een intuïtionist, dat wil zeggen iemand die gelooft dat we de beschikking hebben over een apart ‘zintuig’ dat ons direct kennis over morele waarden verschaft. Zie Mackie 1977: 38.
[138] Minder op de medemens gerichte denkers kiezen wellicht voor de interpretatie ‘een handeling is goed als hij mijn geluk bevordert’. Zie Banning 1972: 127-132 over de ethiek van het egoïsme.
[139] Die uitwerkingen worden respectievelijk daad- en regelutilisme (Hudson 1983: 215-216), of direct en indirect utilisme (Heeger 1985: 35-37), genoemd.
[140] Zie, b.v., Moore 1942; Hudson 1983: hoofdstuk 3.
[141] In zekere zin zou ‘naturalistische fout’ een betere vertaling van ‘naturalistic fallacy’ zijn, het gaat hier niet om een onjuiste redenering of een ten onrechte getrokken conclusie, maar om een interpretatiefout.
[142] Zie Hudson 1983: 100-105.
[143] Het is niet gebruikelijk om deze interpretatie onder het hoofdje ‘naturalisme’ onder te brengen. Ik doe dat hier alleen om de lezer niet met een overdaad aan onderscheidingen te overstelpen.
[144] Hudson 1983: 87.
[145] Hudson 1983: 253.
[146] Hudson 1983: 253-265. Hume’s theorie over moraal kan worden geïnterpreteerd als een poging moraal te beschrijven en te verklaren zonder de verdergaande pretentie morele voorschriften te geven (of het moest zijn het advies je neer te leggen bij wat nu eenmaal zo is). Zie de weergave van Maris 1981: 50-59.
[147] Met name Searl (zie de herdruk van zijn artikel uit 1964 in Foot 1967) en Gewirth. Voor een bespreking van Gewirth’ theorie zie Hudson 1983: 277-294, en de bijdrage van Veldhuis in Hubbeling en Veldhuis 1985: 81-113. Richards’ benadering, die ik verderop bespreek, is – ten dele – door Gewirth’ aanpak geïnspireerd. Zie verder ook Mackie 1977, Maris 1981.
[148] Op zich is ‘ought implies can’ een ethisch principe zonder uiteindelijke rechtvaardiging, of het moest zijn dat het het opstellen van regels waaraan men zich niet houden kan voorkomt. Maar daarmee ga je ervan uit dat het het wenselijke effect van morele principes is dat mensen zich er strikt aan houden. En daar ben ik eerlijk gezegd nog helemaal niet van overtuigd. Ik heb soms een diep respect voor mensen die streven naar iets onmogelijks. Zeker zolang ze dat doen in het besef dat het inderdaad om iets onmogelijks gaat. Een handvol eenvoudige voorbeelden moge hier volstaan: de tien geboden zouden alle tien geschrapt moeten worden als ‘ought’ inderdaad ‘can’ impliceerde. (En in zekere zin is dat ook b.v. Ruse’ conclusie – zie Ruse 1984a.) Maar dat neemt natuurlijk niet weg dat men ervoor kiezen kan dat principe aan te hangen.
[149] Het voorbeeld is van Searl, met één verschil: volgens Searl volgt de plicht tot betalen uit de belofte en de – feitelijke – betekenis van het beloven alleen!
[150] Voor een uitgebreide weergave van Comtes ideeën zie Maris 1981.
[151] Zie Maris 1981.
[152] Wellicht is deze weergave te simpel. Als ik Maris goed begrijp dan had Spencer een instrumentalistische opvatting over het moreel goede: goed is wat genot verschaft. Morele oordelen zijn de uitdrukking van ons zoeken naar genot. Het doel van de moraal acht hij het in stand houden van het leven. Maar dat kan alleen zo zijn als het leven meer genot dan narigheid verschaft. Zo niet dan zou, volgens Spencer, gedrag dat een eind aan het leven maakte goed zijn. Zo bezien is Spencer eerder een hedonistisch descriptivist dan een voorvechter van evolutionaire ethiek. Hij gebruikt de evolutietheorie slechts om af te leiden wat het leven en dus het genot en dus de moraal het beste dienen zal. En daar is niet meer mis mee dan dat het een erg groot vertrouwen in wetenschappelijke theorieën impliceert. Tegelijkertijd echter vat Spencer evolutie op als een ontwikkeling naar steeds hogere (ook in een morele zin) levensvormen, met de mens aan de top. En wie zou zich aan die ontwikkeling willen onttrekken? Die ontwikkeling is trouwens onontkoombaar, dus van een echte ethische keuze is nauwelijks sprake. Zo bezien speelt evolutiebiologie wel een erg grote rol, groter dan die van de feiten die we soms nodig hebben om uit algemene meer specifieke normen af te leiden. En dat rechtvaardigt het vermoeden dat Spencer wel degelijk waarden ontleent aan zijn versie van de evolutietheorie (zie verder Maris 1981: 103-120).
[153] ‘Scientists and humanists should consider together the possibility that the time has come for ethics to be removed temporarily from the hands of the philosophers and biologicized’ (Wilson 1975: 562). Wilson lezen is erg moeilijk, ik geef mijn eigen interpretaties dus graag voor betere.
[154] Evolutionair gezien zeer oude hersenonderdelen.
[155] ‘… the emotional control centers in the hypothalamus and limbic system of the brain flood our consciousness with all the emotions … that are consulted by ethical philosophers who wish to intuit the standards of good and evil. What, are we then compelled to ask, made the hypothalamus and limbic system? They evolved by natural selection. That simple biological statement must be pursued to explain ethics and ethical philosophers, …, at all depths’ (Wilson 1975: 3).
[156] Het komt nog het duidelijkst aan bod in een artikel dat hij samen met de filosoof Ruse geschreven heeft. Zie paragraaf 4.2.2.
[157] Wilson 1978.
[158] ‘... the … dilemma … can be stated as follows: Which of the censors and motivators should be obeyed and which ones might better be curtailed or sublimated? … At some time in the future we will have to decide how human we wish to remain … To chart our destiny means that we must shift from automatic control based on our biological properties to precise steering based on biological knowledge. … only hard-won empirical knowledge of our biological nature will allow us to make optimum choices among the competing criteria of progress’ (Wilson 1978: 6-7).
[159] Zie citaat in vorige noot. ‘Optimaal’ is een goed voorbeeld van een meerplaatsig begrip (zie 1.2.2) dat vaak eenplaatsig wordt gebruikt.
[160] Wilson 1978: 147-153.
[161] Wilson 1978: 130.
[162] Let wel, ik beweer niet dat hij dat bewust doet. Het is waarschijnlijk eerder zo dat hij wat al te onkritisch staat tegenover zijn eigen vooropgezette gevoelens.
[163] Zie bijvoorbeeld Barnett 1980; King 1980; Kitcher 1985; Musschenga 1981; Ruse 1979.
[164] Door af te leiden op welke manier bepaalde gewenste zaken tot stand gebracht kunnen worden, gegeven de manier waarop mensen dankzij hun evolutionaire verleden in elkaar zitten. Volgens Singer 1981: 67-68 kan dat alleen wanneer je werkt met een ethiek geformuleerd in termen van gevolgen van gedrag. Helemaal overtuigend is dat niet. Ook wanneer je gedrag niet beoordeelt op grond van de effecten, maar louter op grond van de motieven, dan zul je toch wellicht kunnen voorspellen onder welke, daardoor moreel wenselijke, omstandigheden bepaalde hoogstaande motieven het best tot ontwikkeling kunnen komen.
[165] Zie bijvoorbeeld Campbell 1978; Singer 1981; Murphy 1982; Ruse 1984a; Ruse en Wilson 1985; Richards 19863 en 1986b.
[166] Murphy 1982.
[167] ‘At some point in a chain of justification one simply digs in one’s heels and stops with something (happiness perhaps) that is just accepted as a value, not proven to be one. But if this is so, what guarantee do we have that this value … is not simply arbitrary and thus all values based on it … arbitrary? The ‘whys’, it seems, logically outlast all possible answers – a comprehensive skepticism that one might well find terribly disquieting’ (Murphy 1982: 12).
[168] ‘... for if a religious value such as ‘doing God’s will’ will not guarantee meaningfulness, it is hard to see how some secular value … will fare any better. Such appeals will also be open to the same logical problem – namely a dependency upon postulated values that lie beyond the possibility of rational proof. The upshot, then, is simply this: Nothing works; and thus there is one very real sense in which human life is absurd. The things that matter to us the most really cannot be proven to be true’ (Murphy 1982: 1617).
[169] ‘Our unjustly maligned ‘animal nature’ – our passions and patterns of evolved habitual behavior keep us together through shared values and commitments and will continue to do so even at the loss of a covering intellectual rationale’ (Murphy 1982: 19).
[170] ‘Now there are many ways (some of them obscure, some of them quite clever) in which one might attempt to save religion from this onslaught of scientific discovery: interpret religion metaphorically or symbolically, talk of faith and mystery instead of evidence and reason, insist that religion is mainly ethics (‘ultimate concern’) and not metaphysics, regard religion and science as two radically different ways of talking about the world (language games or forms of life) that are compatible since we talk for different purposes in the two cases, or even use Humean skepticism on behalf of religion – i.e., recall that Hume mounted a powerful skeptical case against induction (the basic methodology of science) and thus showed that the person who rejects science cannot be proven irrational for so doing. … The above, of course, are all various strategies of avoidance’ (Murphy 1982: 33).
[171] Zie Ruse en Wilson 1985. Voor een uitgebreidere versie van hetzelfde verhaal zie Ruse 1984a.
[172] Sloep (persoonlijke mededeling) merkte hier terecht op dat échte superhersenen natuurlijk heel snel kunnen beslissen. Het bezwaar van Ruse en Wilson geldt dus alleen langzame superhersenen (en dat lijkt een contradictie).
[173] ‘In short, to make us altruistic in the adaptive, biological sense, our biology makes us altruistic in the more conventionally understood sense of acting on deeply held beliefs about right and wrong’ (Ruse en Wilson 1985: 51).
[174] Zie hoofdstuk 3 en bijlage 1.
[175] ‘Does the sociobiological scenario just sketched justify the same moral code that religionists believe to be decreed by God? Or that some philosophers believe to exist apart from humanity, like a mathematical theorem?’ (Ruse en Wilson 1985: 51).
[176] ‘... morality will usually function best when it is done genuinely, because we think it right. Humans who act for noble motives are more efficient than those always looking for returns’ (Ruse 1984a: 179).
‘Even though our powers of reason show us there is no objective morality, as humans each and every one of us feels that there is. Even the most hardline sociobiologist conducts his or her daily life as though there were such objectivity’ (Ruse 1980: 192).
‘In speaking thus of illusion, we are not saying that ethics is nothing, and should now be thought of as purely dreamlike. …, ethics is a shared illusion of the human race. If this were not so it would not work’ (Ruse en Wilson 1985: 52).
[177] Ruse en Wilson 1985: 52. Zie ook Ruse 1980: 185-191.
[178] Om een simpel en bekend voorbeeld te geven: een beest dat vier jagers in een bosje ziet verdwijnen en er even later weer twee uit ziet komen zou, als hij in plaats van 2+2=4 geloofde dat 2+0=4, denken dat het bosje vrij van jagers is. En dat kan hem zijn leven kosten!
[179] Campbell 1978.
[180] Hij noemt dat een ‘scientific mediational normative ethics’ (Campbell 1978: 68).
[181] ‘Scientific mediational normative ethics could be developed for any ultimate goal, but such development is obviously of little social or practical moral use unless the ultimate goals to which they are directed enjoy considerable popular consensus. Here evolutionary theory can enter in a reflexive way. From the neo-Darwinian version of evolutionary theory it would follow that all organisms, including humans, have as a build-in goal the survival of their own genes in future generations. But we can no doubt expand the set of goals. I feel certain that it includes more than the future survival of the human genome.’ (Campbell 1978: 69).
[182] Lastiger wordt het natuurlijk als anti-abortionisten zich niet op hun eigen plicht tegenover hun eigen kind beroepen, maar op het recht van elk ongeboren kind. Het conflict dat dan ontstaat betreft in zekere zin de vraag of het recht op abortus een kwestie van persoonlijke ethiek is of een kwestie van sociale ethiek. En ik zie niet hoe Campbells eerste waarde in dat conflict een oplossing kan bieden.
[183] Campbell bedoelt waarschijnlijk de menselijke genenpoel.
[184] Volgens sommigen begaat bijvoorbeeld Mill deze misstap. Op een gegeven moment lijkt hij uit het feit dat ieder streeft naar zijn eigen geluk af te leiden dat we dus allemaal streven naar ons gemeenschappelijke geluk. Maar er is een verschil: Mills woorden lijken ook anders uitgelegd te kunnen worden (zie Mackie 1977), bij Campbell kan dat niet.
[185] Zie Ruse 1979: 203 en Kitcher 1985: 430-431.
[186] Murphy 1982: 112, noot 21.
[187] Zie Richards 1986a en 1986b.
[188] ‘Some versions of evolutionary ethics have represented the current state of our society as ethically sanctioned. Other versions … have identified long-term trends in evolution, which they ipse facto deem good;… But [Richards’ theorie] … prescribes neither of these alternatives. It does not specify a particular social arrangement as being best; rather, it supposes that men will seek the arrangement that appears best to enhance the community good. The conception of what constitutes such an ideal pattern will change through time and over different cultures’ (Richards 1986a: 280).
[189] Zoals bijvoorbeeld Thomas 1986: 317-320.
[190] ‘rv supposes that a moral sense has evolved in the human group. ‘Moral sense’ names a set of innate dispositions that, in appropriate circumstances, move the individual to act in specific ways for the good of the community. … These specific sympathetic responses and pricks to action together constitute the core of the altruistic attitude’ (Richards 1986a: 272).
[191] ‘…‘to justify' means 'to demonstrate that a proposition or system of propositions conforms to a set of acceptable rules, a set of acceptable factual propositions, or a set of acceptable practices'. If this is an accurate rendering of the concept of justification, then the justification of first moral principles and inference rules must ultimately lead to an appeal to the beliefs and practices of men, which of course is an empirical appeal. So moral principles ultimately can be justified only by facts’ (Richards 1986a: 285-286).
[192] Tenzij we hem ervan kunnen overtuigen dat ‘morele’ mensen óns oordeel delen en het zijne niet. Maar dat zal niet meevallen. Alleen voor een moreel relativist is het echt vanzelfsprekend dat iemand met wie je het in ethische kwesties volslagen oneens bent toch een moreel hoogstaand mens kan zijn. En voor ethisch relativisten is deze rechtvaardigingspoging van Richards van meet af aan al onaanvaardbaar.
[193] De veronderstelling dat ‘… moral impulses are part of the equipment of every normal human being … can easily degenerate into the position that those who do not share our particular moral outlook are for that reason subhuman’ (Trigg 1986: 334).
[194] ‘In reference to structured contexts, ‘ought to occur’, ‘ought to be’, ‘ought to act’, etc. typically mean ‘must occur,’ ‘must be,’ ‘must act, provided there is no interference’. Structured contexts involve causal processes. Typically, ‘ought’ adds to ‘must’ the idea that perchance some other cause might disrupt the process …
In the context of the evolutionary constitution of human moral behavior, ‘ought’ means that the person must act altruistically, provided he has assessed the situation correctly and a surge of jealousy, hatred, greed, etc. does not interfere’ (Richards 1986a:290).
[195] Wilson 1978.
[196] Ruse l984b.
[197] Het meest geruchtmakende alternatief op dit moment is een theorie – als hij die naam verdient – die zichzelf als ‘wetenschappelijk creationisme’ afficheert. Philip Kitcher 1982 veegt daar op klaarblijkelijk overtuigende wijze de vloer mee aan, en dat gebeurt eveneens, zij het binnen een heel ander filosofisch kader, in het boek van Houtman c.s. 1986. Een creationist binnen mijn eigen kennissenkring wierp tegen dat beide boeken zijn gebaseerd op creationistische geschriften waar veel creationisten zelf ook afstand van hebben genomen. Ik kan dat niet beoordelen. Op dit moment biedt het verhaal van de evolutiebiologie de meest verhelderende kijk op ontstaan en ontwikkeling van het leven op aarde. Ik houd graag de mogelijkheid open dat dat oordeel berust op onvoldoende kennis van andere literatuur.
[198] De ‘just-so-stories’ waar de paleontoloog Gould 1980 het over heeft.
[199] Ik heb natuurlijk een grenzeloos vertrouwen in mijn eigen gezonde verstand, maar dat van anderen vertrouw ik alleen maar als het klopt met het mijne. En helaas komt het vaak voor dat mensen door hun gezonde verstand geheel verschillende kanten worden opgestuurd. Dat is een belangrijke oorzaak van verschillen in oordeel over veel sociobiologische verhalen. Het gaat daarbij vrijwel altijd om evolutionaire verhalen waarvan we de feitelijke vooronderstellingen niet kunnen toetsen, bijvoorbeeld omdat gedrag niet fossiliseert.
[200] Dat was zo ruwweg bij de overgang van Mioceen naar Plioceen. Over de precieze datering daarvan verschilt men van mening. Volgens Leaky en Lewin 1978 was dat 5 miljoen jaar, volgens Morgan 1972 zo’n 15 miljoen jaar, en volgens McAlester 1968 7 miljoen jaar geleden. De onenigheid kan ’m zitten in de naamgeving van periodes, over de ouderdom van verschillende fossielen bestaat minder verschil van mening.
[201] Zie bijvoorbeeld Morris 1967.
[202] Zie Morgan 1972. De geïnteresseerde lezer verwijs ik graag nog naar haar latere boek uit 1982, dat uitsluitend en veel uitgebreider over de theorie van ons waterige verleden gaat. Het andere hoofdthema in Morgan 1972, de masculinistische vooroordelen, ontbreekt hierin.
[203] Als we ervan uitgaan dat mannen wat vaker aan land kwamen dan vrouwen zouden we zo zelfs het verschil in lichaamsbeharing kunnen verklaren. En eveneens het feit dat vrouwen als lange-afstandszwemmers mannen ruimschoots evenaren (zie Wilson 1978: 132).
[204] Tanner 1981.
[205] Zihlman 1984 betoogt zeer uitgebreid dat de dwergchimpansee (Pan paniscus (die inmiddels veelal met de uit het Engelse taalgebied overgewaaide naam bonobo wordt aangeduid; toevoeging 2009)) een nog iets beter model is dan de gewone chimpansee (Pan troglodytes).
[206] Een aardig detail in dit verband is dat het verschil in lichaamsgrootte tussen mannen en vrouwen volgens traditioneel georienteerde biologen vrij groot is, volgens b.v. Wilson 1978 zo’n 20 tot 30%, terwijl Tanner het over een procent of vijf heeft. Dat lijkt met elkaar in strijd, maar na wat rekenwerk blijkt het toch nog aardig te kloppen. Wilson kijkt naar lichaamsgewicht, en Tanner naar lichaamslengte. En als je even aanneemt dat lichaamsgewichten zich ruwweg verhouden als derde-machten van de lichaamslengte dan is een eerste oorzaak van het verschil al duidelijk. Wie 5% langer wordt, zou bij behoud van lichaamsverhoudingen bijna 16% zwaarder worden. Bovendien neemt Wilson de vrouw als uitgangspunt, stelt haar op 100%, en berekent dan hoeveel van zulke procenten de man zwaarder is. Tanner doet het omgekeerde, zij stelt de man op 100% en berekent hoeveel van die procenten de vrouw korter is. Wilson heeft daardoor meer procenten nodig voor hetzelfde verschil.
[207] Hoewel de Australopithecus-soort met de zwaarste kaken en kiezen, de overduidelijk herbivore A. boisei, door niemand meer als een voorloper van de mens gezien wordt, is de gangbare opvatting onder palaeontologen toch dat ook onze australopithecine voorouders vooral planteneters waren.
[208] Leaky en Lewin 1978.
[209] Hun commentaar betreft een andere publicatie van Tanner dan die (uit 1981) waarop de vorige paragraaf gebaseerd is, maar is hierop evenzeer van toepassing.
[210] Vooral Elaine Morgan zal niet door iedereen als autoriteit worden beschouwd. Ze maakte carrière als tv-producente en journaliste. Ik denk dat ze zeer goed weet waar ze het over heeft, maar om critici die aan academische papieren hechten ter wille te zijn zal ik haar verder niet in de vergelijking betrekken.
[211] Zie Wilson 1978: 130.
[212] Zie bijvoorbeeld Sayers 1982: 58.
[213] Trivers 1974 en 1985.
[214] Wilson 1978: 128-154.
[215] Meulenbelt 1984: 73 e.v.; Sayers 1982: 58.
[216] Wilson 1978.
[217] Zie Von Denffer et al 1967: 207-208.
[218] Ghiselin 1966; Hull 1978.
[219] Een taxon, i.t.t. de categorie. Zie Ruse 1973: 122.
[220] Zie Van der Steen en Voorzanger 1986.
[221] Ten overvloede: het gaat hier maar om een voorbeeld. Het is heel wel denkbaar dat juist de tweede persoon minder terughoudend zou reageren.
[222] Edel en Edel, 1986: 45; Sahlins 1977.
[223] ‘Among the Trobriand islanders of the South Pacific a basic code of mutual obligation is so ingrained that a man will abandon a day’s highly paid pearl diving for a puzzled white trader in order to do a day’s fishing for an inland neighbor in return for a few yams or other farm produce, because the obligations to a trading partner must always be fulfilled. But their neighbors from the nearby island of Dobu who also live by a complicated system of trade relations, consider any cheating one can actually get away with as a mark of the greatest shrewdness, an admired steppingstone to success. … Anthropologists have found that differences go far beyond these matters of specific rules and regulations, or even goals and values. They reach into any and every part of the phenomena and structure of morality’ (Edel en Edel 1968: 21).
[224] Zie bijvoorbeeld: Wilson 1978; Barash 1979; Van den Berghe 1980.
[225] Dat mechanisme trekt zich er niets van aan of de samen opgroeiende kinderen biologisch gezien nu wel of niet verwant zijn. Ook tussen kinderen die samen in één leeftijdsgroep in een kibboets opgroeien zijn huwelijken schaars (zie Wilson 1978: 37).
[226] Hier valt natuurlijk wel meer over te zeggen. In een samenleving waarin nauwe verwanten wel regelmatig huwelijken aangaan, zullen ongunstige recessieve genen ten gevolge van natuurlijke selectie waarschijnlijk verdwijnen. Dat bezwaar geldt dus maar tijdelijk. Uiteindelijk draait het, bij het verklaren van incest- (liever: inteelt-) vermijding, allemaal om de vraag wat de functie is van genetische diversiteit, en hoe groot die idealiter zou moeten zijn. Daarover zijn biologen het alles behalve eens (Zie Trivers 1985: 315 e.v.).
[227] Deze samenvatting is gebaseerd op de inleiding van Lumsden en Wilson 1981.
[228] De lezer zij er zich van bewust dat ‘mind’ in het engels iets minder hoogdravend en etherisch klinkt dan ‘geest’ in het Nederlands.
[229] Ik neem aan dat ze de drie primaire kleuren plus het licht/donkeronderscheid bedoelen.
[230] Ik doe m’n best zo netjes mogelijk weer te geven wat de auteurs beweren. Soms dwingt me dat iets op te schrijven waarvan ik eigenlijk niet weet wat het betekent.
[231] Waaruit dat blijkt blijft wat duister, maar de auteurs zijn hier zeer beslist over.
[232] Het oordeel, intussen, van sommigen dergenen die deze modellenbouwerij wel kunnen volgen is behoorlijk vernietigend (zie bijvoorbeeld Kitcher 1985 en Maynard Smith 1982b).
[233] Ik neem hier de wellicht laakbare gewoonte over om over dé sociale wetenschappen te spreken, alsof er niet een enorme verscheidenheid aan sociaal-wetenschappelijke disciplines (en stromingen binnen elke discipline) zou zijn. De lezer bedenke dat ik me daarvan bewust ben, en dat ik de voor de hand liggende nuanceringen alleen weglaat om hem onleesbaar lange zinnen te besparen.
[234] Als we bij evolutie ten minste niet meteen denken aan de tientallen miljoenen jaren durende ontwikkeling van boomkruinbewonende zoogdiertjes met een muisachtig voorkomen tot de mens zoals we die nu om ons heen zien. Ik neem aan dat sociale wetenschappers die het belang van genetische factoren voor de verschillen tussen culturen ontkennen dat belang niet zullen ontkennen als het gaat om bijvoorbeeld de overgang van Homo erectus naar Homo sapiens.
[235] Onderzoek naar de genetische bepaaldheid van menselijk gedrag staat in de kinderschoenen, en het is de vraag of het daar ooit aan zal kunnen ontgroeien. Experimenteel onderzoek is, op ethische gronden, zo goed als uitgesloten, en we mogen hopen dat dat zo blijven zal, maar zonder experimenteel onderzoek valt er weinig definitief vast te stellen. Natuurlijk is er onderzoek, en Lumsden en Wilson citeren dat uitvoerig, waaruit blijkt dat de kans op het voorkomen van bepaalde ‘gedragingen’ en voorkeuren afhangt van het voorkomen daarvan bij ouders, en er is uitgebreid onderzoek gedaan naar de overeenkomsten tussen eeneiige tweelingen en tussen tweeeiige tweelingen waaruit blijkt dat eeneiige tweelingen onderling meer op elkaar lijken dan tweeëiige tweelingen. Maar dat betekent allemaal niet meer dan dat er aanwijzingen zijn dat verschillen in gedrag tussen mensen binnen één gemeenschap ten dele genetisch bepaald zouden kunnen zijn. Waar het om gaat in de controverse die Lumsden en Wilson hier wat ongelukkig aandikken is of er gedragsverschillen zijn tussen verschillende culturen die voortkomen uit systematische genetische verschillen tussen mensen uit verschillende culturele gemeenschappen. Ik heb de indruk dat daarvoor niet veel aanwijzingen zijn.
[236] Zie Cavalli-Sforza en Feldman 1981.
[237] Cavalli-Sforza en Feldman 1981: 341-344.
[238] Denk bijvoorbeeld aan heterosis, het geval waarin de heterozygoot de hoogste fitness heeft en daardoor het voortbestaan (of wellicht beter: het in elke generatie opnieuw ontstaan) van zelfs een lethale homozygoot kan veroorzaken. Voor bijvoorbeeld Dawkins (1976, 1982) is zo’n uitzondering reden om selectie geheel op het niveau van ‘losse’ genen te definiëren. Op zich leidt dat tot een fraai en schijnbaar uiterst consistent theoretisch bouwwerk. Maar er bestaat twijfel aan de verklarende kracht van Dawkins’ benadering (zie b.v. Sober en Lewontin 1982). Voor een bespreking van meningsverschillen over selectieniveaus zie Van der Steen en Voorzanger 1984a.
[239] Een vooronderstelling die in veel van de door de auteurs genoemde gevallen bepaald niet vanzelf spreekt! Het besnijden van jongetjes valt op hygiënische gronden heel wel te verdedigen, en voorzover veel en vet eten de gezondheid schaadt zou dat best tot de dood kunnen leiden in een fase waarin dat voor het nageslacht economisch voordeel oplevert (erfenis; geen zorg voor hulpbehoevende bejaarden). Ik zie de zin niet zo van dat soort profijtanalyses, maar als men ze maakt laat men dat dan ten minste goed doen.